
INTRODUCCIÓN
En la costa atlántica de México, los bancos ostrícolas están compuestos principalmente por el ostión americano Crassostrea virginica, esta especie se distribuye desde Tamaulipas hasta Yucatán y habita comúnmente en lagunas y esteros (García-Cubas y Reguero, 2007). Para Veracruz, representa un recurso pesquero importante y es una de las más productivas del país, donde, para el 2013 se estimó que este Estado incorporó alrededor del 45 % (19 422 ton) de la producción nacional ostrícola (42 945 ton), particularmente la laguna de Pueblo Viejo, Tamiahua y Tampamachoco fueron las de mayor aporte (Baqueiro-Cárdenas et al., 2013).
Los bancos ostrícolas de C. virginica son importantes ecológicamente para las lagunas costeras, dado que proveen alimento y hábitat para una amplia variedad de peces e invertebrados de importancia económica y ecológica (Peterson y Lipcius, 2003; Susan-Tepetlan y Aldana-Aranda, 2008; Blomberg et al., 2018), además, las valvas ofrecen un sustrato apropiado para la fauna vágil y para la fijación de una diversa comunidad epibionte (Tolley y Volety, 2005; Stunz et al., 2010; Blomberg et al., 2018). No obstante, este tipo de conocimiento es escaso para el suroeste del golfo de México, contados estudios solo resaltan la importancia de los bancos ostrícolas para el asentamiento de diversos grupos de invertebrados (Salazar-Vallejo y González, 1990; Ruíz y López-Portillo, 2006).
Para la costa atlántica mexicana, al menos dos estudios aportan información de la macrofauna asociada a los bancos ostrícolas. Primero, destaca el estudio realizado por Susan-Tepetlan y Aldana-Aranda, (2008) quienes identificaron y cuantificaron la macrofauna en bancos ostrícolas para las lagunas Carmen, Machona y Mecoacán en Tabasco, y concluyen que moluscos y crustáceos son los principales grupos abundantes. Mientras, Coutiño-Rodríguez (1982), reporta para los bancos ostrícolas de la laguna La Mancha en Veracruz, un ensamblaje faunístico más diverso representado por moluscos, crustáceos, poliquetos, esponjas, briozoarios y platelmintos. Aunado a lo anterior, resulta necesario realizar más estudios que determinen la estructura comunitaria que se asocia a los bancos ostrícolas, por ello, la presente investigación contribuye con información del ensamblaje faunístico en bancos ostrícolas de tres lagunas costeras del norte de Veracruz, México.
OBJETIVOS
- Determinar la composición, abundancia y similitud de la macrofauna asociada los bancos ostrícolas de Crassostrea virginica, en las lagunas de Pueblo Viejo (PV), Tamiahua (TAH) y Tampamachoco (TAP), Veracruz, México.
MATERIALES Y MÉTODOS
Área de estudio. El presente trabajo se realizó en tres lagunas costeras ubicadas al norte del Estado de Veracruz, México. La laguna de Pueblo Viejo (PV), situada en el municipio de Villa Cuauhtémoc se localiza a 22° 05’ y 22° 13’ N y 97° 50’ y 97° 57’ W (Fig. 1), presenta una superficie de 93.7 km2, tiene una extensión de 15 km a lo largo en sentido norte-sur y a lo ancho 9.5 km aproximadamente (Contreras-Espinosa, 1985; Lara-Domínguez et al., 2011), la profundidad de este ecosistema es de 1.5 m, con rango de temperatura de 26.6-30.4 °C y salinidad de 11.9-27.3 ups (Contreras-Espinosa, 1993). Laguna de Tamiahua (TAH) se extiende entre los paralelos 21° 15´ y 22° 06’ N y 97° 23’ y 97° 46’ W (Fig. 1), es la más grande en superficie con 880 km2 y presenta una longitud aproximada de 85 km y una anchura máxima de 18 km (Contreras-Espinosa, 1993; De la Lanza-Espino y Cáceres-Martínez, 1994), con una profundidad de 2-3 m, la temperatura fluctúa de 21-31 °C y con variaciones de salinidad de 16-37 ups (Contreras-Espinosa, 1993). Laguna de Tampamachoco (TAP), localizada en la llanura costera a 9 km al noroeste de la ciudad de Tuxpan; se ubica en las coordenadas 20° 18’ y 21° 02’ N y 97° 19’ y 97° 22’ W (Fig. 1), presenta una longitud y anchura de 10.6 km y 2.7 km respectivamente, ocupando una superficie de 15 km2aproximadamente (Reguero y García-Cubas, 1991; Contreras y Castañeda, 2004), la profundidad es de 1.5 m, con variaciones de temperatura de 18.3-32.1 °C y salinidad de 15.3-36.6 ups (Contreras-Espinosa, 1993).
Para cada laguna se seleccionaron tres bancos ostrícolas de C. virginica considerando como criterio los de mayor producción de acuerdo al conocimiento de los pescadores (Fig. 1), los cuales se mencionan a continuación: Santa Clara (22º 09´ N y 97º 51´ O), El Ciruelo (22º 09´N y 97º 52´ O) e Isleta Grande (22º 09´N y 97º 54´ O) para PV; Cabecillas (21° 18´ N y 97° 26´ O),
La Palma (21° 09´ N y 97° 22´ O) y Panzacola (21° 21´ N y 97° 27´O) para TAH; Pipiloya (21° 01´ N y 97° 21´ O), Restinga de Afuera (21° 00´ N y 97° 21´ O) y El Tubo (21° 00´ y 97° 21´ O) para TAP. En cada banco ostrícola se realizaron muestreos mensuales (julio-diciembre, 2016) para determinar la estructura comunitaria de la macrofauna asociada a los bancos de C. virginica y realizar comparaciones a nivel mensual, así como también indagar similitudes entre lagunas costeras.
Fase de campo. Para la recolecta de organismos se utilizó la metodología descrita por Baqueiro et al. (2013) y Ascencio et al. (2016) que consistió en utilizar el arte de pesca conocido como “gafas” la cual se compone de dos rastrillos, unidos, formando una pinza; se manipularon desde la borda de la lancha introduciéndolas abiertas rascando el banco a muestrear, al cerrarlas, se obtienen los ostiones y epibiontes asociados a las ostras. En cada banco ostrícola, se realizaron varias extracciones hasta obtener 60 ostiones con tallas igual o mayor a 40 mm de longitud total, de tal manera que se recolectaron mensualmente 180 ostiones para cada laguna. Por lo tanto, englobando las tres lagunas, mensualmente fueron recolectados 540 ostiones. Los ostiones elegidos se colocaron en una caja de plástico de 60 cm de largo y 40 cm de alto y se trasladaron al laboratorio de Biotecnología Ambiental de la Facultad de Ciencias Biológicas y Agropecuarias, Tuxpan, Veracruz.
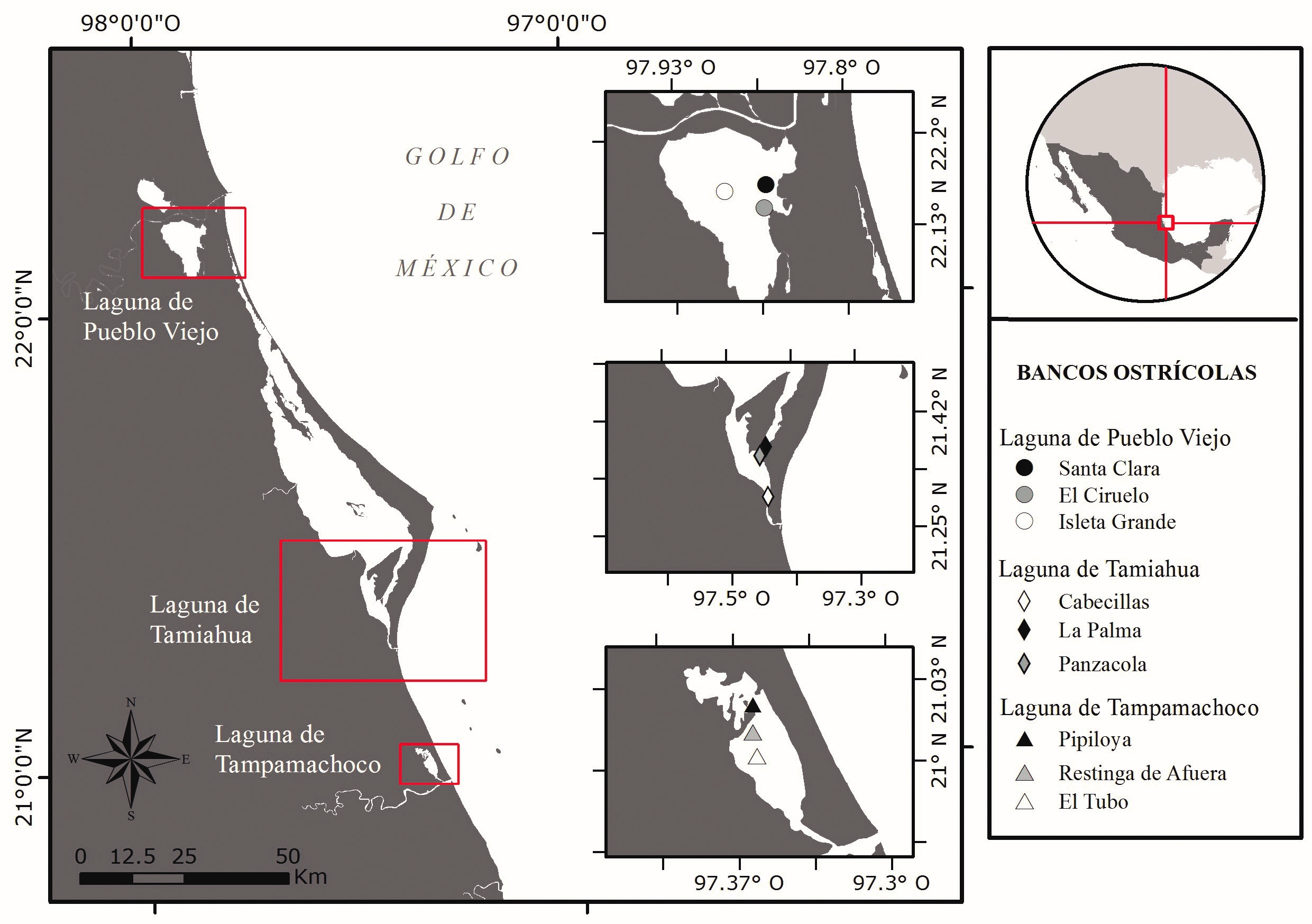
Figura 1. Localización geográfica de los bancos ostrícolas (Crassostrea virginica) de las lagunas de Pueblo Viejo, Tamiahua y Tampamachoco, Veracruz, México.
Fase de laboratorio. Los organismos asociados a las valvas de C. virginica, se separaron con la ayuda de un desconchador y un cuchillo, posteriormente se depositaron en un frasco de plástico transparente con capacidad de 1.5 L y se fijaron en solución de formol al 10 %. Todos los organismos fueron identificados empleando claves taxonómicas y guías de identificación especializadas para cada grupo zoológico (Verril, 1892; Hyman, 1951; Moore y López, 1970; García-Cubas y Reguero, 2004, 2007). Posterior a la identificación taxonómica, los organismos fueron contabilizados al nivel más próximo a especie.
Cabe mencionar que se realizó una revisión bibliográfica para clasificar a los organismos por su tipo de asociación (epibionte, parásito, móvil y sedentario) y los hábitos alimentarios (filtrador, carnívoro, omnívoro, herbívoro, micrófago, ramoneador) de las especies identificadas utilizando las obras de García-Cubas y Reguero (2004, 2007), De León-González et al. (2009). Además se consultó la World Register of Marine Species (www.marineespecies.org) para actualizar la nomenclatura de los nombres científicos.
Con la información obtenida (presencia y abundancia) se evaluó la eficiencia de muestreo para cada laguna con base en el estimador no paramétrico Chao 1. Con la riqueza estimada se calculó la representatividad del muestreo (%) dividiendo el valor final de la riqueza observada (multiplicado por 100) entre el valor final de la riqueza esperada. El desempeño del estimador se evaluó calculando el sesgo y exactitud con base en las fórmulas que detallan Walther y Moore (2005). Los valores de la riqueza observada y estimada se obtuvieron con el programa EstimateS (usando 100 aleatorizaciones) versión 9 (Colwell, 2013).
También se utilizó el modelo asintótico de Clench para predecir el número de especies existentes en cada laguna, así también para evaluar la calidad del inventario y estimar el esfuerzo de muestreo realizado, todo este proceso se realizó con base en el trabajo de Jiménez-Valverde y Hortal (2003). Para cada laguna se elaboraron curvas acumulativas de la riqueza observada y del estimador Chao 1 y Clench para fines comparativos.
También se estimaron índices ecológicos basados en la abundancia: riqueza específica, diversidad de Shannon, equitatividad y diversidad verdadera de orden 1 (1D), este índice se obtuvo calculando el exponente de la diversidad de Shannon (1D = exp H’), dicho índice expresa el número efectivo de especies que tendría una comunidad integrada por especies igualmente abundantes, excluyen especies raras y dominantes, tiene la ventaja de realizar comparaciones de la diversidad verdadera de dos o más comunidades (Moreno, 2001). Cada parámetro ecológico fue estimado a nivel de laguna y a nivel mensual, además se determinaron diferencias significativas entre los meses de muestreo de cada laguna estudiada mediante la prueba de Kruskal-Wallis dado que los datos no cumplieron los supuestos de normalidad y homogeneidad, para ello se utilizó el programa Statistica 7 (StatSoft, 2004).
Las semejanzas entre los bancos ostrícolas con base en la composición y abundancia de la macrofauna asociada, se determinaron con el método de escalamiento multidimensional no métrico (NMDS) utilizando el índice de similitud de Bray-Curtis, este método de ordenación representó gráficamente los resultados en dos dimensiones las proximidades (grupos afín) de las unidades de muestreo. Así mismo, se utilizó la prueba de similitud ANOSIM para averiguar diferencias significativas en los ensamblajes faunísticos asociados a los bancos ostrícolas. También se realizó un análisis de similitud porcentual (SIMPER) para determinar cuáles especies contribuyeron en la similitud de los grupos más parecidos. Estos análisis multivariantes se realizaron con el programa Primer Versión 7 (Clarke y Gorley, 2015).
RESULTADOS
Se recolectó un total de 1 606 organismos macrobentónicos asociados a los bancos ostrícolas de C. virginica en las lagunas de PV, TAH y TAP, en las que se identificaron seis phyla, 18 ordenes, 24 familias y 28 especies (Tabla I). Siendo Mollusca y Annelida los de mayor número de especies con 10 y siete respectivamente. Al menos 15 especies de la macrofauna asociada a C. virginica se registraron en los tres sistemas lagunares, mientras las especies registradas en un solo sistema lagunar fueron Vitrinella floridana para la laguna PV; Hydroides dianthus, Tanais dulongii, Eurypanopeus depressus y Panopeus lacustris fueron observadas en TAP (Tabla I).
En la laguna PV se registraron 17 especies macrofaunísticas asociadas a los bancos ostrícolas, cifra que es cercana a la predicción del estimador Chao 1 de 17.5 especies, lo que significa que se registró el 97.1 % de la macrofauna. Así también, el modelo de Clench estimó 17.04 especies (a = 28.46548; b = 1.665824), lo que reafirma un esfuerzo de muestreo eficiente (99.4 %). De manera comparativa, las especies observadas y estimadas presentan tendencia a la asíntota (Fig. 2A). Los grupos faunísticos con mayor riqueza de especies fueron Mollusca, Arthropoda y Annelida con siete, cinco y cuatro especies respectivamente, los demás grupos taxonómicos están representados por una especie (Fig. 3A).
Tabla I. Listado sistemático de la macrofauna béntica asociada a bancos ostrícolas (Crassostrea virginica) de las lagunas de Pueblo Viejo (PV), Tamiahua (TAH) y Tampamacho (TAP), Veracruz, México

Para los bancos ostrícolas de la laguna de TAH, la composición macrofaunística asociada fue de 22 especies, el estimador Chao 1 presentó un buen desempeño al estimar la misma cantidad de especies, mientras la predicción del modelo de Clench fue 23.4 especies (a = 29.4017258; b = 1.25969), esto representa un 94 % de macrofauna registrada en los bancos ostrícolas. Para esta laguna, la curva acumulativa de especies observadas y estimadas se muestran asintóticas (Fig. 2B). Los invertebrados de mayor riqueza de especies fueron Mollusca (9 spp.) y Annelida (6 spp.), seguido de Arthropoda (4 spp.), los restantes invertebrados presentaron solo una especie (Fig. 3A).
Respecto a los bancos ostrícolas de la laguna TAP, se registró mayor riqueza macrofaunística asociada, encontrándose 27 especies, esta cifra coincide con la predicción del estimador Chao 1. Así mismo, el modelo de Clench estimó 27.5 especies (a = 149.1885; b = 5.420398), lo que significa que se registró el 98 % de la riqueza faunística asociada a los bancos ostrícolas. Estos resultados se visualizan en las curvas acumulativas de especies observadas y estimadas, las cuales se muestran asintóticas (Fig. 2C). De igual manera, Mollusca, Arthropoda y Annelida fueron los de mayor riqueza de especies, mientras los grupos con menor número de taxa fueron Porifera, Platyhelminthes y Bryozoa (Fig. 3A).
En cuanto a la abundancia, en la laguna PV, se cuantificó un total de 382 especímenes asociados a C. virginica, los moluscos fueron los más abundantes, seguido de los anélidos (Fig. 3B). Brachiodontes exutus fue la especie mayor representada con 121 individuos, seguido de Neritina usnea con 37 y Marphysa sanguinea con 33 individuos, el resto de las especies presentaron abundancias <30 especímenes (Fig. 4A). En la laguna de TAH, la abundancia registrada fue de 367 especímenes, de igual manera, moluscos y anélidos fueron los más representados con 149 y 122 individuos (Fig. 3B). No obstante, a nivel de especie, la de mayor abundancia fue Nereis oligohalina y Stylochus ellipticus con 76 y 35 organismos contabilizados, el resto de las especies presentaron abundancias <31 individuos (Fig. 4B). Para la laguna TAP se cuantificó la mayor cantidad de macroinvertebrados (857 individuos) asociados a C. virginica, los más abundantes fueron los artrópodos con 293 individuos, seguido de los moluscos con 288 individuos (Fig. 3B). Por consiguiente, Amphibalanus eburneus fue la más numerosa con 121 individuos, seguido de T. dulongii con 73 especímenes, las restantes especies presentaron <61 individuos (Fig. 4C).
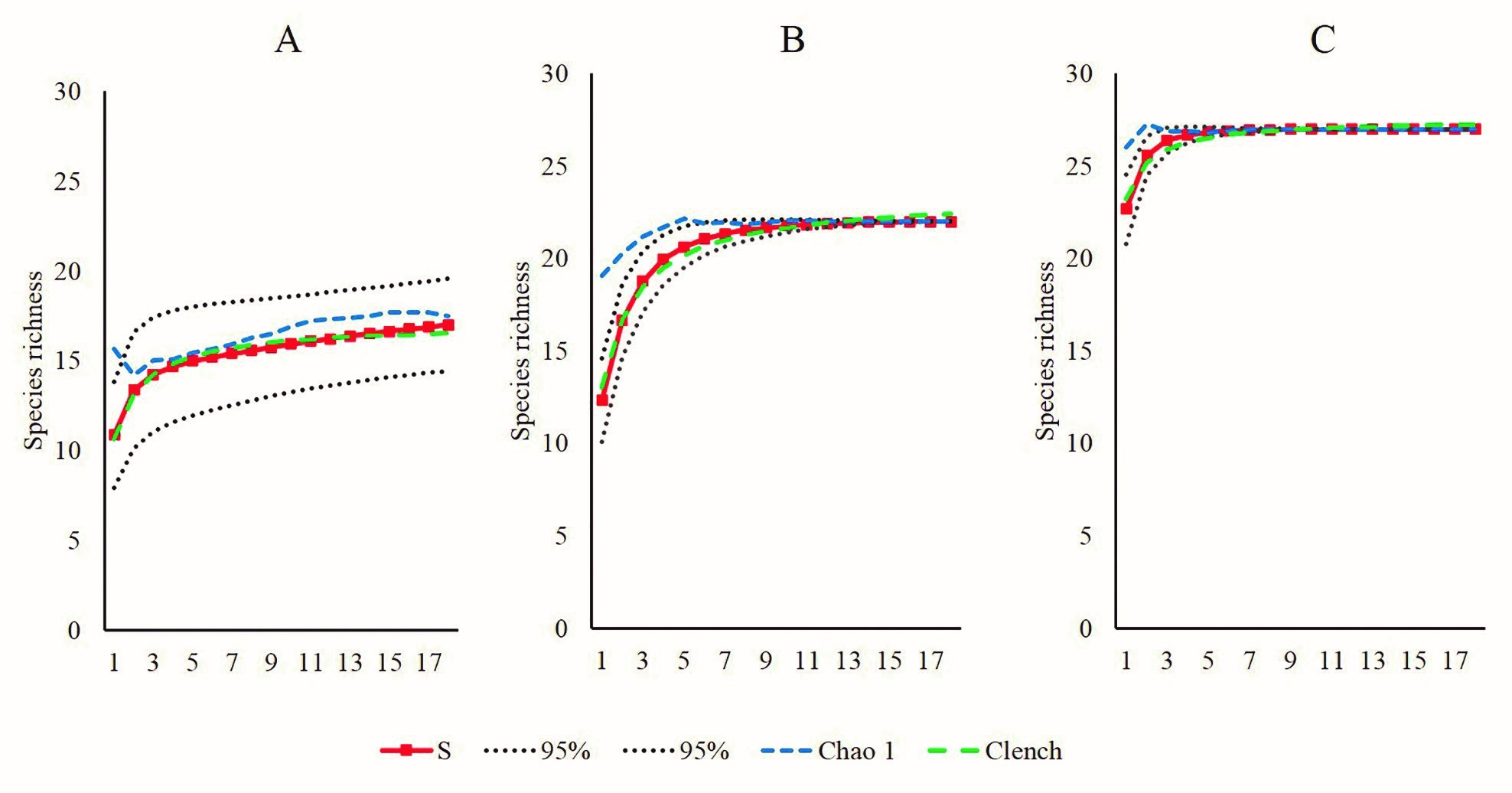
Figura 2. Curvas de acumulación de especies observadas y estimadas. A, laguna Pueblo Viejo; B, Tamiahua; C, Tampamachoco.
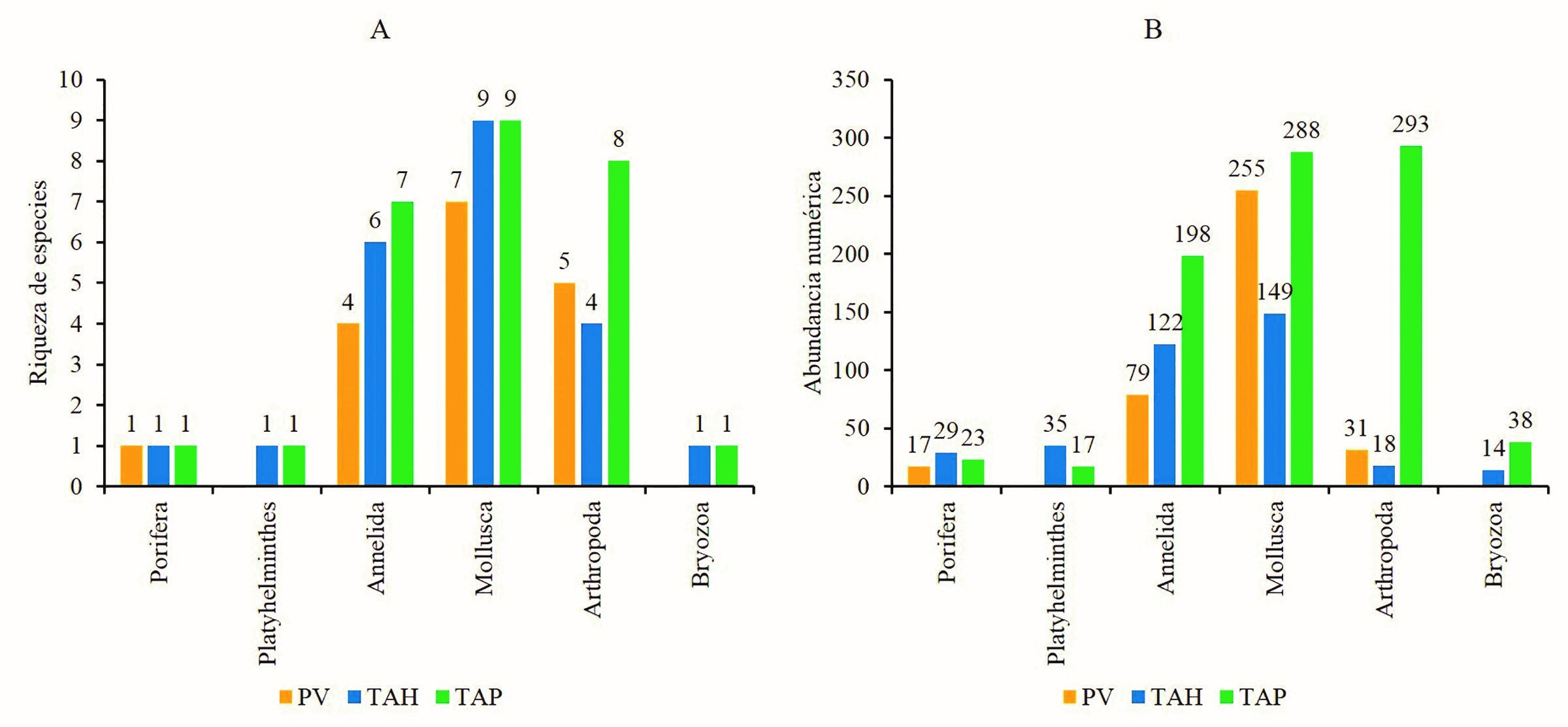
Figura 3. Riqueza y abundancia de la macrofauna bentónica asociada a bancos ostrícolas (Crassostrea virginica) de las lagunas de Pueblo Viejo (PV), Tamiahua (TAH) y Tampamachoco (TAP), Veracruz, México. A, riqueza; B, abundancia.
Por otra parte, la mayoría de la macrofauna encontrada en los bancos ostrícolas fueron epibiontes y móviles, mientras que los menos representados en abundancia fueron las formas parásitas y de vida sedentaria (Fig. 5A). Así también, la mayoría fueron de hábitos filtradores, seguido de los carnívoros y omnívoros (Fig. 5B). De las tres lagunas costeras, en TAP se registró mayor abundancia de macrofauna epibionte y móvil, así mismo, la mayoría fueron de hábitos filtradores (Fig. 5A, B).

Figura 4. Curvas de abundancia de las especies más representativas asociadas a los bancos ostrícolas (Crassostrea virginica). A, Pueblo Viejo; B, Tamiahua; C, Tampamachoco.
Con relación al análisis mensual, los bancos ostrícolas de la laguna PV presentaron una riqueza de 13 especies en julio y un máximo de 15 especies en los meses posteriores, excepto diciembre (Fig. 6A), no obstante, agosto y noviembre fueron los meses con mayor número de especímenes recolectados, mientras julio y septiembre presentaron menor abundancia (Fig. 6B).
Para la laguna de TAH, septiembre y diciembre se registraron 17 taxa, mientras en noviembre se registró mayor riqueza con 20 especies (Fig. 6A), en cambio, la mayor abundancia fue registrada en julio, posteriormente decreció en los demás meses con abundancias <70 individuos (Fig. 6B). Para los bancos ostrícolas de la laguna TAP, fue el ecosistema que presentó mayor riqueza y abundancia macrofaunística, siendo noviembre el mes con mayor número de taxa (27 especies) y diciembre como el mes con mayor abundancia con 162 individuos (Fig. 6B).

Figura 5. Valores de abundancia de la macrofauna asociada a los bancos ostrícolas (Crassostrea virginica) de las lagunas de Pueblo Viejo (PV), Tamiahua (TAH) y Tampamachoco (TAP), Veracruz, México. A, hábitos de vida; B, tipo de nutrición.
En general, la estructura comunitaria de la macrofauna asociada a los bancos ostrícolas es equitativa, dado que se registraron valores superiores a J = 0.81 (Fig. 6C). De manera específica, el ensamblaje macrofaunístico asociado a los bancos ostrícolas de la laguna PV mostró un valor mínimo de J = 0.81 en agosto, mientras en julio se registró un valor máximo de J = 0.91 (Fig. 6C), sin embargo, estos contrastes no fueron significativos (H = 4.04094; P = 0.543537). En los bancos ostrícolas de la laguna de TAH, la comunidad macrofaunística asociada presentó valores >0.85 de equitatividad (Fig. 6C), siendo agosto, noviembre y diciembre los meses que presentaron los valores más altos (J = 0.93), por lo que estadísticamente no se detectaron diferencias significativas entre los meses de estudio (H = 10.3313; P = 0.066375). De igual manera, la macrofauna asociada a los bancos ostrícolas de la laguna TAP exhibió valores >0.9 de equitatividad (Fig. 6C), en agosto y septiembre se registró el valor más bajo (J = 0.91) y el valor máximo se registró en julio y octubre (J = 0.95), no obstante, estos contrastes no son estadísticamente significativos (H = 4.1345; P = 0.530218). Con relación a la diversidad verdadera, se observa una misma tendencia, existiendo mayor cantidad de especies efectivas en los meses estudiados para la laguna TAP, contrariamente en PV, todos los meses registraron valores de diversidad verdadera menor a 10 especies efectivas (Fig. 6D).
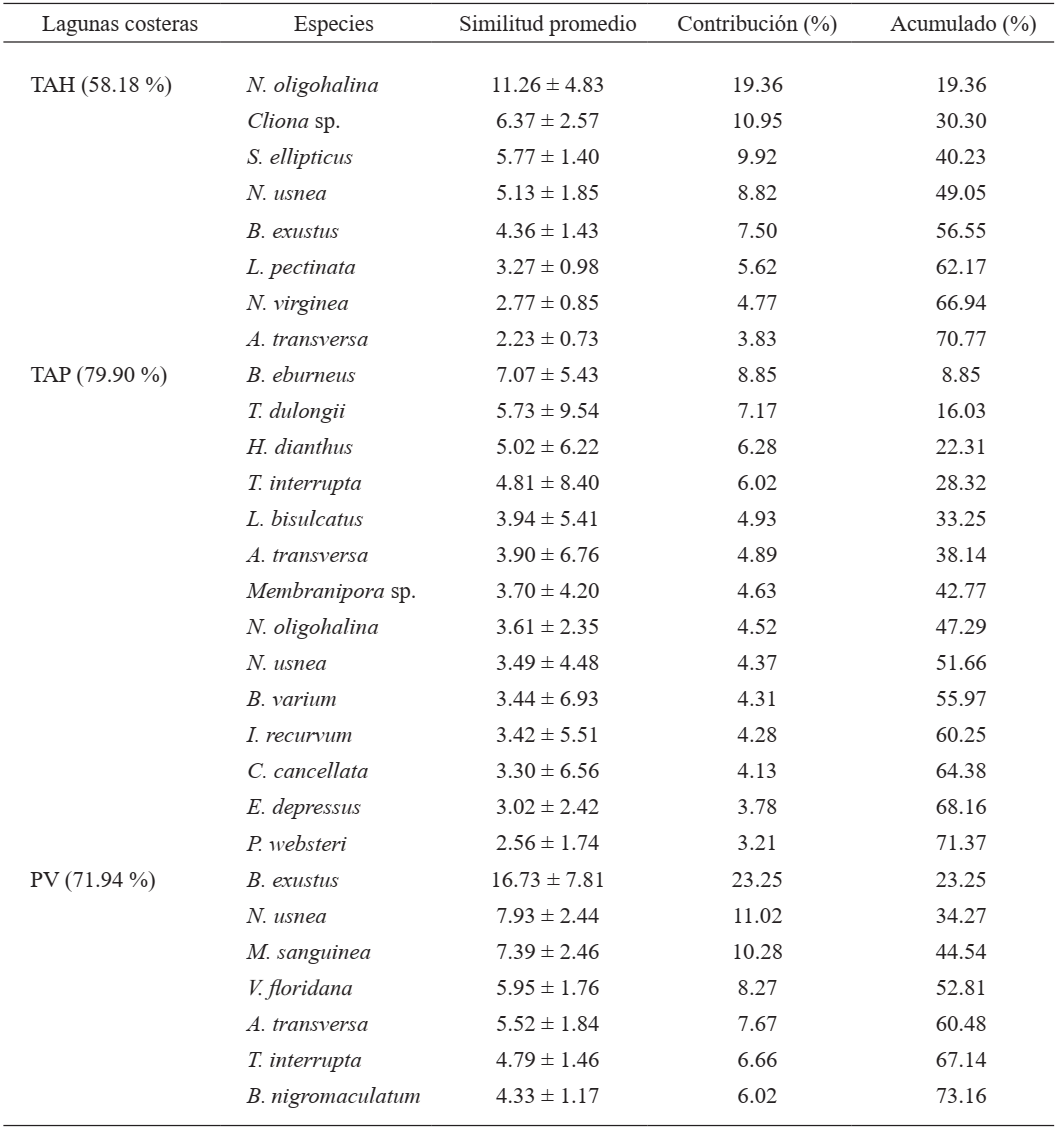
Por otra parte, los bancos ostrícolas de PV, TAH y TAP presentaron diferencias significativas con base en la composición y abundancia de la macrofauna asociada de acuerdo al análisis ANOSIM (R global = 0.897, P <0.001). Estos contrastes se confirman con el análisis NDMS, donde se muestra la separación de las lagunas costeras (Fig. 7). Con base en el análisis SIMPER, 14 especies contribuyeron en la similitud de TAP (79.9 %), así también, siete especies favorecieron la similitud de PV (71.94 %) y seis especies para TAH (58.18 %), (Tabla II).

Figura 6. Valores mensuales de riqueza, abundancia, equitatividad y diversidad verdadera de la macrofauna bentónica asociada a bancos ostrícolas (Crassostrea virginica) en las lagunas de Pueblo Viejo (PV), Tamiahua (TAH) y Tampamachoco (TAP), Veracruz, México. A, riqueza; B, abundancia; C, equitatividad; D, diversidad verdadera.
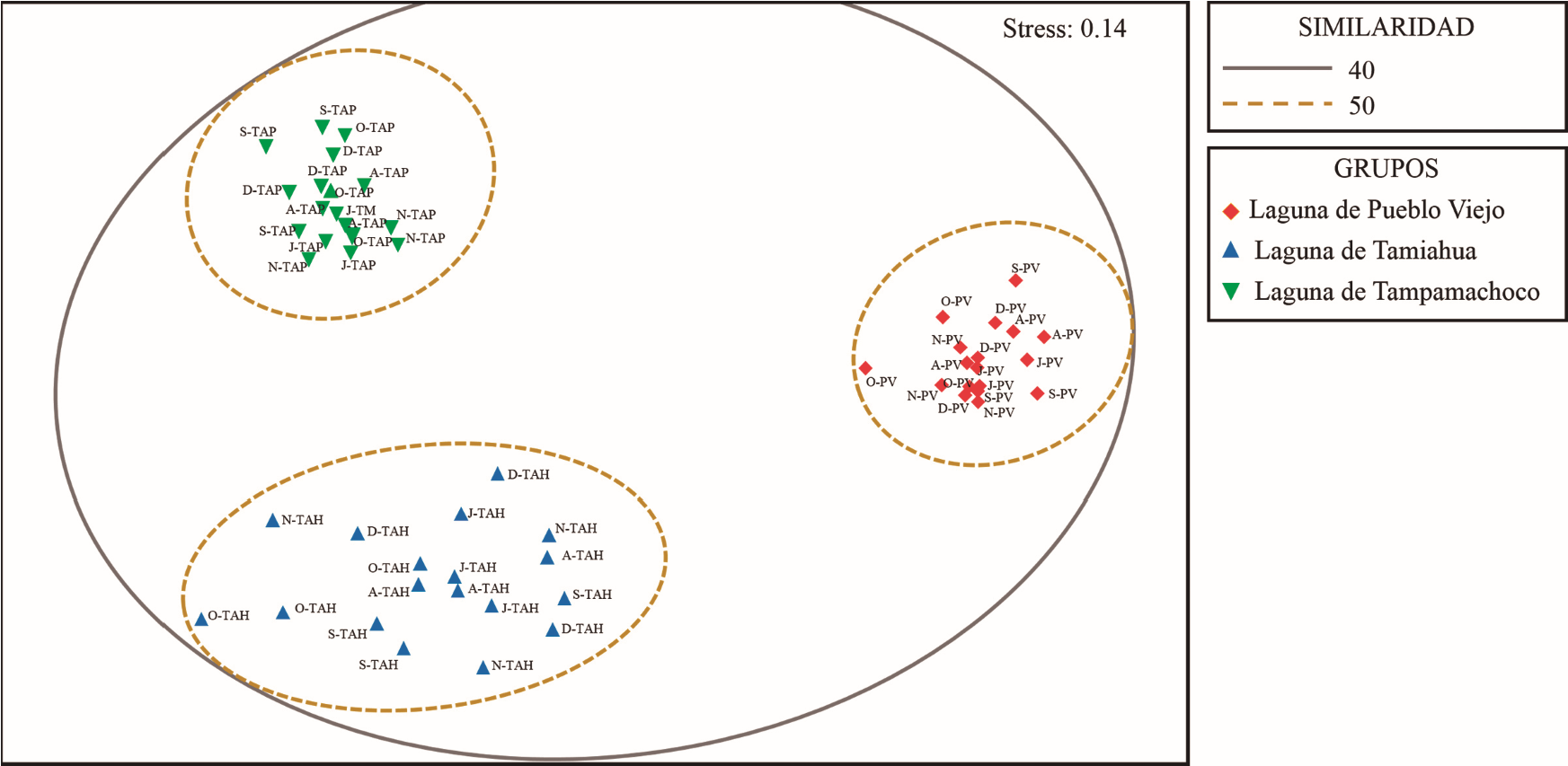
Figura 7. Análisis de escalamiento multidimensional no métrico (nMDS) realizado a los bancos ostrícolas de la laguna Pueblo Viejo (PV), Tamiahua (TH) y Tampamachoco (TM).
Tabla II. Valores de similitud porcentual (SIMPER) de las especies macrofaunísticas que contribuyeron en la semejanza de los bancos ostrícolas de las lagunas costeras Pueblo Viejo (PV), Tamiahua (TAH) y Tampamachoco (TAP), Veracruz, México
DISCUSIÓN
Los resultados del presente estudio indican que los bancos ostrícolas de las lagunas Pueblo Viejo, Tamiahua y Tampamachoco constituyen un importante sustrato biogénico que es aprovechado por invertebrados sésiles y vágiles. La macrofauna encontrada en los bancos ostrícolas está compuesto por esponjas, anélidos, moluscos, crustáceos y briozoarios, estos resultados coinciden a los ensamblajes faunísticos que se reportan para otras regiones del atlántico (Tolley et al., 2005; Tolley y Volety, 2005; Blomberg et al., 2018).
Por otra parte, la macrofauna de moluscos y crustáceos encontrada (17 especies) en el estudio es menor a la que se reporta para otros ecosistemas similares de la costa atlántica mexicana, dado que en la laguna Carmen-Machona, Tabasco, se han registrado 26 especies de moluscos y 12 de crustáceos (Susan-Tepetlan y Aldana-Aranda, 2008). Así también, para los bancos ostrícolas de la laguna La Mancha, Veracruz, se ha reportado una riqueza macrofaunística compuesta por 12 especies de moluscos, 10 de crustáceos, cuatro de poliquetos y tres especies de peces (Coutiño-Rodríguez, 1982).
De las tres lagunas costeras estudiadas, los bancos ostrícolas de PV presentaron menor riqueza de especies, lo cual se puede atribuir a que tienen una menor extensión superficial, sin embargo, los bancos de ostión de TAP resultaron ser los de mayor riqueza macrofaunística a pesar que la laguna de TAH es la de mayor tamaño, por ello es necesario investigar si las características fisicoquímicas como la salinidad y la naturaleza del sustrato de cada laguna influyen en estos contrastes, o bien si dependen de la tasa de crecimiento y longitud de los ostiones, debido a que C. virginica es de mayor tamaño en la laguna de TAP (Vidal-Briseño et al., 2015).
Dentro del ensamblaje macrofaunístico que se establece en los bancos ostrícolas, es importante destacar que se identificaron varias especies perforadoras y parásitas, por ejemplo: Cliona sp., esta esponja perfora las valvas de C. virginica y se observó en todos los bancos ostrícolas. Se ha documentado que miembros de este género son capaces de afectar las tasas de crecimiento y provocar la mortalidad de los ostiones (Carver et al., 2010; Carroll et al., 2015). Entre los parásitos encontrados fueron S. ellipticus en TAP y TAH, este platelminto es un activo depredador del ostión (Landers y Rhodes, 1970) y se tiene un antecedente histórico para la laguna de TAH en la sequía de 1988, se registró una proliferación de estos organismos provocando la mortalidad masiva de los ostiones (DOF, 2014). Este platelminto se ha reportado también para el sistema de lagunas Carmen-Machona, Tabasco (Gómez, 1980).
Asimismo, se observó a Polydora websteri en las lagunas TAH y TAP, es un parásito con actividad erosionadora y se ha notificado que causa problemas en el crecimiento de los ostiones (Haigler, 1969), haciéndolos vulnerables a enfermedades, lo cual podría conducir a problemas en el rendimiento de la producción de los bancos ostrícolas y mortalidad en algunos casos (Wargo y Ford, 1993; Martin y Britayev, 1998; Cáceres-Martínez y Vásquez-Yeomans, 2013).
De igual manera, crustáceos parásitos fueron registrados como Zaops ostreum, el cual se caracteriza por depredar los bancos ostrícolas, causando problemas fisiológicos en los ostiones juveniles (Hanke et al., 2015). También se observó a T. dulongii, esta especie se encontró en el interior de los ostiones, lo cual acontece a través de la etapa larval (Byers et al., 2014). Es importante señalar la presencia de E. depresssus, este decápodo se registró en TAP y se caracteriza por ser un depredador de ostiones juveniles (Kulp et al., 2011).
La mayoría del ensamblaje macrofaunístico que se establece en los bancos ostrícolas son epibiontes, representados por bivalvos y balanos, los cuales fueron los más abundantes en los tres sistemas lagunares, resultados que coinciden a lo que se reporta para ecosistemas similares (Tolley y Volety, 2005; Tolley et al., 2005; Susan-Tepetlan y Aldana-Aranda, 2008; Hanke et al., 2015; Blomberg et al., 2018). Esto sugiere que balanos y bivalvos son los grupos más exitosos en los bancos ostrícolas, lo cual se puede atribuir a que tienen la capacidad de fijarse a las valvas del ostión por la secreción de cemento y de un biso (García-Cubas y Reguero, 2007), así también, son tolerantes a amplios rangos de salinidad y de temperatura (Bacon, 1971; Henry y McLaughlin, 1975; García-Cubas y Reguero, 2007), presentan altas tasas de reproducción y crecimiento e. g. A. eburneus (El-Komy y Kajihara, 1991), inclusive habitualmente cohabitan y obtienen los mismos recursos alimenticios filtrando partículas suspendidas de la columna del agua (García-Cubas y Reguero, 2007).
Los bancos de ostión americano, también son fundamentales para la fauna móvil, ya que fue el segundo grupo con mayor abundancia, lo cual puede estar relacionado proporcionalmente a una mayor presencia de epibiontes, tal como se registró en TAP y TAH. Por ello, la abundancia de carnívoros y omnívoros móviles en los bancos ostrícolas se puede atribuir por propósitos de alimentación (Susan-Tepetlan y Aldana-Aranda, 2008), fungiendo como depredadores (e. g. S. ellipticus, P. websteri, Z. ostreum) de los epibiontes y de los propios ostiones.
Las especies más representativas en abundancia fueron distintas en cada banco ostrícola. En la laguna PV, la más numerosa fue B. exustus, es epibionte de las valvas del ostión y adopta hábitos filtradores, le sigue N. usnea, de hábitos sedentarios y micrófagos, ambas especies son típicas del sistema lagunar (Reguero y García-Cubas, 1993) y presentan amplia tolerancia a la salinidad (García-Cubas y Reguero, 2004). Por el contrario, en la laguna de TAH, las más representativas fueron carnívoros como N. oligohalina, seguido del parásito S. ellipticus, este resultado sugiere que los bancos ostrícolas pueden presentar depredación por estos invertebrados móviles. Para la laguna TAP, los bancos ostrícolas están sujetos a mayor colonización de epibiontes filtradores ya que los más abundantes fueron justamente A. eburneus, seguido de H. dianthus, también fue importante la abundancia de crustáceos móviles como T. dulongii.
Solamente A. eburneus y E. depressus, son citados de igual manera como abundantes para los arrecifes de ostión de las lagunas Carmen-Machona y Mecoacán (Susan-Tepetlan y Aldana-Aranda, 2008). En estos sistemas lagunares las especies dominantes fueron Ischadium recurvum, Crepidula plana, A. eburneus, Amphibalanus improvisus y E. depressus (Susan-Tepetlan y Aldana-Aranda, 2008). En bancos ostrícolas artificiales también sitúan a A. eburneus como especie dominante (Meyer y Towsend, 2000). La macrofauna epibionte también se ha estudiado en las raíces de R. mangle, sin embargo, el ensamblaje macrofaunístico es distinto a la de los bancos ostrícolas dado que en la laguna de TAP y la Mancha, las especies más abundantes en las raíces del mangle rojo son Amphibalanus sp., Crassostrea rhizophorae, I. recurvum y Mytilopsis leucophaeata (Ruiz y López-Portillo, 2014; Lucas y De la Cruz-Francisco, 2018).
En general, la comunidad macrofaunística de los bancos ostrícolas es equitativa, lo cual se puede atribuir a la abundancia de especies epibiontes que cohabitan compartiendo los mismos recursos como el espacio y alimento, obteniendo los recursos alimenticios de la columna del agua, misma que es abundante por la alta tasa de sedimentación que presentan las lagunas costeras (Contreras-Espinosa, 1993; Calva y Torres-Alvarado, 2000). En consecuencia, esto favorece un ambiente uniforme y diverso en la estructura comunitaria.
Aunado a lo anterior, la equitatividad es mayor en los bancos ostrícolas con mayor riqueza y abundancia de epibiontes y depredadores como se observó en la laguna TAP, mientras la equitatividad es menor en ambientes con menor diversidad y abundancia de especies como la registrada en PV. La equitatividad también se ha reportado para otras comunidades estuarinas de la laguna de TAP, principalmente con la macrofauna que se asocia a las raíces de R. mangle (Lucas y De la Cruz-Francisco, 2018), así mismo con la comunidad de moluscos de las lagunas TAP (Reguero y García-Cubas, 1991) y PV (Reguero y García-Cubas, 1993).
El análisis de similitud evidencia que los bancos ostrícolas de cada laguna son afines entre sí con relación a la composición y abundancia de la macrofauna asociada, pero son significativamente diferentes al compararlas entre las tres lagunas costeras. Esto se atribuye a que el ensamblaje macrofaunístico asociado a los bancos ostrícolas de PV presentaron menor riqueza y abundancia, donde la especie típica fue un epibionte filtrador: B. exustus. Mientras, en TAP, la macrofauna asociada en los bancos ostrícolas fueron más diversos en especies y en abundancia, representados por A. eburneus y H. dianthus, ambos de hábitos filtradores. Contrariamente, en TAH, los bancos ostrícolas son caracterizados principalmente por carnívoros (N. oligohalina y S. ellipticus), seguido de epibiontes filtradores. Todo esto parece indicar que los modos de vida y los grupos tróficos parecen ser determinantes en la estructura comunitaria de la macrofauna que se asocia a los bancos ostrícolas de C. virginica.
CONCLUSIONES
El ensamblaje faunístico identificado en los bancos ostrícolas de las lagunas del norte de Veracruz (PV, TAH y TAP) corrobora la importancia que desempeñan en los ecosistemas costeros, ya que proveen un sustrato biogénico idóneo para el asentamiento de macroinvertebrados bentónicos que fungen como residentes (epibiontes) y transitorios (fauna móvil). No obstante, la estructura macrofaunística asociada a los bancos ostrícolas es distinta en las lagunas estudiadas, por lo que es necesario que posteriores estudios relacionen los atributos comunitarios con variables ambientales como las características físico-químicas, la profundidad y la extensión de los bancos ostrícolas, estos factores podrían explicar la dinámica ecológica de estos ensamblajes macrofaunísticos.
AGRADECIMIENTOS
A Yehimy Yunery Martínez Hernández por su ayuda en el trabajo de campo y procesamiento de datos. También agradecemos a Antonio Leija Tristán del Depto. Ecología, Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, por el apoyo en la identificación de las especies.
Literatura Citada
Ascencio, L. A., M. Enríquez, I. Martínez y D. Aldana. 2016. Efecto de la temperatura y la salinidad en el ciclo reproductor de hembras y machos de Crassostrea virginica (Bivalvia: Ostreidae). Revista de Biología Tropical, 64: 449-459.
Bacon, P. R. 1971. The maintenance of a resident population of Balanus eburneus (Gould) in relation to salinity fluctuation in a Trinidad mangrove swamp. Journal of Experimental Marine Biology and Ecology, 6: 187-198.
Baqueiro-Cardenas, E., J. Bello-Pineda, I. Martínez-Morales, P. San Martín-Del Ángel, I. Hernández-Herrera, F. Lango-Reynoso, M. Enríquez-Díaz y D. Aldana-Aranda. 2013. Estrategias poblacionales del ostión americano Crassostrea virginica del Estado de Veracruz, (19-30 pp.). En: Aldana-Aranda, D., M. Enríquez-Díaz y E. V Elías-Gutiérrez (Eds.), Manejo de los recursos pesqueros de la cuenca del Golfo de México y del mar Caribe, Universidad Veracruzana, Xalapa,
Blomberg, B. N., T. A. Palmer, P. A. Montagna y J. B. Pollack. 2018. Habitat assessment of a restored oyster reef in South Texas. Ecological Engineering, 122: 48-61.
Byers, J. E., T. L. Rogers, J. H. Grabowski, A. R. Hughes, M. F. Piehler y D. L. Kimbro. 2014. Host and parasite recruitment correlated at a regional scale. Oecologia, 174: 731-738.
Cáceres-Martínez, J. y R. Vásquez-Yeomans. 2013. Enfermedades, parásitos y episodios de mortalidad de ostiones de importancia comercial en México y sus implicaciones para la producción. Ciencia Pesquera, 21: 5-48.
Calva, B. L. G. y R. Torres-Alvarado. 2000. Distribución de carbohidratos, carbono y nitrógeno orgánico en sedimentos de tres lagunas costeras del golfo de México. Hidrobiológica, 10 (2): 101-114.
Carroll, J. M., K. A. O’Shaughnessy, G. A. Diedrich y C. M. Finelli. 2015. Are oyster being bored to death? Influence of Cliona celata on Crassostrea virginica condition, growth and survival. Diseases of Aquatic Organisms, 117: 31-44.
Carver, C. E., I. Thériault y A. L. Mallet. 2010. Infection of cultured eastern oysters Crassostrea virginica by the boring sponge Cliona celata, with emphasis on sponge life history and mitigation strategies. Journal of Shellfish Research, 29 (4): 905-915.
Clarke, K. R. y R. N. Gorley. 2015. Getting started with PRIMER V7. PRIMER-E: Plymouth. Plymouth Marine Laboratory, 20.
Colwell, R. K. 2013. EstimateS, Version 9.1: Statistical estimation of species richness and shared species from samples. Storrs: University of Connecticut, United States of America.
Contreras-Espinosa, F. 1985. Las lagunas Costeras Mexicanas. México: Centro de Ecodesarrollo, Secretaría de Pesca, 253 pp.
Contreras-Espinosa, F. 1993. Ecosistemas Costeros Mexicanos. México: Comisión Nacional para el Conocimiento y uso de la Biodiversidad, Universidad Autónoma Metropolitana, 415 pp.
Contreras, F. y O. Castañeda. 2004. La biodiversidad de las lagunas costeras. Ciencias, 076: 46-56.
Coutiño-Rodríguez, A. 1982. Contribución al conocimiento de la fauna acompañante de la población ostrícola de la Laguna de La Mancha, Veracruz. Tesis licenciatura. Universidad Veracruzana, 85 pp.
De la Lanza-Espino, G. y C. Cáceres-Martínez. 1994. Lagunas costeras y el litoral mexicano:
geología (pp. 13-74). En: de la Lanza-Espino, G. y C. Cáceres-Martinez (Eds), Lagunas Costeras y el Litoral Mexicano. UNAM-UABCS.
De León-González, J. A., J. R. Bastida-Zavala, L. F. Carrera-Parra, M. E. García-Garza, A. PeñaRivera, S. I. Salazar-Vallejo y V. Solís-Weiss. 2009. Poliquetos (Annelida: Polychaeta) de México y América Tropical. México: Universidad Autónoma de Nuevo León, Monterrey, 737 pp.
Diario Oficial de la Federación (DOF). 2014. Plan de Manejo Pesquero de Lisa (Mugil cephalus) y Lebrancha (Mugil curema), en las costas de Tamaulipas y Veracruz. Diario Oficial de la Federación, 31 de marzo de 2014. https://www.inapesca.gob.mx/portal/documentos (Accesado: 14 de abril de 2019).
El-Komy, M. M. y T. Kajihara. 1991. Breeding and moulting of barnacles under rearing conditions. Marine Biology, 108: 83-89.
García-Cubas, A. y M. Reguero. 2004. Catálogo ilustrado de moluscos gasterópodos del Golfo de México y mar Caribe. México: Universidad Nacional Autónoma de México, 157 pp.
García-Cubas, A. y M. Reguero. 2007. Catálogo ilustrado de moluscos Bivalvos del Golfo de México y mar Caribe. México: Universidad Nacional Autónoma de México, 94 pp.
Gómez, A. S. 1980. Frecuencia de Stylochus ellipticus Girard, 1850 (Turbellaria Polycladia), en Crassostrea virginica Gmelin, de lagunas costeras del sur del Golfo de México (1977-1979). Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 51 (1): 1-9.
Haigler, S. A. 1969. Boring mechanis of Polydora websteri inhabiting Crassostrea virginica. American Zoologist, 9: 821-828.
Hanke, M. H., J. M. Hargrove, T. D. Alphin y M. H. Posey. 2015. Oyster utilization and host variation of the oyster pea crab (Zaops ostreum). Journal of Shellfish Research, 34 (2): 281-287.
Henry, D. P. y P. A. McLaughlin. 1975. The barnacles of the Balanus amphitrite complex (Cirripedia, Thoracica). Zoologische Verhandelingen, 141: 1-254.
Hyman, L. H. 1951. The Invertebrates: Platyhelminthes and Rhynchocoela the acoelomate bilateria. EE. UU: McGraw- Hill Book Company, 550 pp.
Jiménez-Valverde, A. y J. Hortal. 2003. Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología, 8: 151-161.
Kulp, R. E., V. Politano, H. A. Lane, S. A. Lombardi y K. T. Paynter. 2011. Predation of juvenile Crassotrea virginica by two species of mud crabs found in the Chesapeake Bay. Journal of Shellfish Research, 30: 1-6.
Landers, W. y E. W. Rhodes. 1970. Some factors influencing predation by the flatworm Stylochus ellipiticus (Girard), on oysters. Chesapeake Science, 11: 55-60.
Lara-Domínguez, A. L., F. Contreras-Espinosa, O. Castañeda-López, E. Barba-Macías y M. A. Pérez-Hernández. 2011. Lagunas Costeras y Estuarios (301-317 pp). En: Cruz-Angón A. (Ed.), La Biodiversidad en Veracruz: Estudio del Estado México: CONABIO, Universidad Veracruzana, Xalapa.
Lucas, M. E. y V. De la Cruz-Francisco. 2018. Macroflora y macrofauna asociada a las raíces sumergidas de Rhizophora mangle (Rhizophoraceae), en la laguna Tampamachoco, Veracruz, México. Revista Colombiana de Ciencia Animal, 10 (1): 31-42.
Martin, D. y T. A. Britayev. 1998. Symbiotic Polychaetes: Review of known species. Oceanography and Marine Biology, 36: 217-340.
Meyer, D. L. y E. C. Townsend. 2000. Faunal utilization of created intertidal eastern oyster (Crassostrea virginica) reefs in the southeastern United States. Estuaries, 23: 34-45.
Moore, H. B. y N. N. López. 1970. A contribution to the ecology of the lamellibranch Dosonia elegans. Bulletin of Marine Science, 20 (4): 980-986.
Moreno, C. E. 2001. Manual de métodos para medir la biodiversidad. México: Universidad Veracruzana (Textos universitarios), 84 pp.
Peterson, C. H. y R. N. Lipcius. 2003. Conceptual progress towards predicting quantitative ecosystem benefits of ecological restorations. Marine Ecology Progress Series, 264: 297-307.
Reguero, M. y A. García-Cubas. 1991. Moluscos de la laguna Tampamachoco, Veracruz, México: Sistemática y Ecología. Anales del Instituto de Ciencias del Mar y Limnología, 18 (2): 289-328.
Reguero, M. y A. García-Cubas. 1993. Moluscos de la laguna Pueblo Viejo, Veracruz, México: Sistemática y Ecología. Anales del Instituto de Ciencias del Mar y Limnología, 20 (1): 77-104.
Ruiz, G. M. y J. López-Portillo. 2006. Los invertebrados. En: Moreno-Casasola, P. (Ed.) Entornos Veracruzanos: la Costa de La Mancha (pp. 341-362). México: Instituto de Ecología A.C., Xalapa, Veracruz.
Ruiz, M. y J. López-Portillo. 2014. Variación espacio-temporal de la comunidad de macroinvertebrados epibiontes en las raíces del mangle rojo Rhizophora mangle (Rhizophoraceae) en la laguna costera de La Mancha, Veracruz, México. Revista de Biología Tropical, 62 (4): 1309-1330.
Salazar-Vallejo, S. I. y N. E. González. 1990. Ecología costera en la región de la Mancha, Veracruz. La ciencia y el hombre, 6: 101-120.
StatSoft. 1984-2004. Statistica for Windows (Software-System for Data-analyses), version 7. USA: Tulsa.
Stunz, G. W., T. J. Minello y L. P. Rozas. 2010. Relative value of oyster reef as habitat for estuarine nekton in Galveston Bay, Texas. Marine Ecology Progress Series, 406: 147-159.
Susan-Tepetlan, P. V. y D. Aldana-Aranda. 2008. Macrofauna bentónica asociada a bancos ostrícolas en las lagunas costeras Carmen, Machona y Mecoacán, Tabasco, México. Revista de Biología Tropical, 56: 127-137.
Tolley, S. G. y A. K. Volety. 2005. The role of oyster in habitat use of oyster reefs by resident fishes and decapod crustaceans. Journal of Shellfish Research, 24 (4): 1007-1012.
Tolley, S. G., A. K. Volety y M. Savarese. 2005. Influence of salinity on the habitat use of oyster reefs in three southwest Florida estuaries. Journal of Shellfish Research, 24 (1): 127-137.
Verrill, A. E. 1892. The marine nemerteans of New England and adjacent waters. Transactions of the Connecticut Academy of Arts and Sciences, 8: 382-456.
Vidal-Briseño, C. I., R. I. Hernández-Herrera, G. Galindo-Cortés y P. San Martín-del Angel. 2015. Estructura poblacional de la captura del ostión Crassostrea virginica en las lagunas de Tamiahua y Tampamachoco, Veracruz, México. Hidrobiológica, 25 (2): 265-273.
Walther, B. A. y J. L. Moore. 2005. The concepts of bias, precision and accuracy, and their use in testing the performance of species richness estimator, with a literature review of estimator performance. Ecography, 28: 815-829.
Wargo, R. N. y S. E. Ford. 1993. The effect of shell infestation by Polydora sp. and infection by Haplosporidium nelsoni (MSX) on the tissue condition of oysters, Crassostrea virginica. Estuaries, 16: 229-234.