
INTRODUCTION
Among the birds that provide the most information for changes in bird community through time, pigeons and doves occupy a special place in Caribbean avifauna. First, the Caribbean currently hosts some twenty Columbidae species (Raffaele et al., 2003), making this family well represented in the region. Second, members of this family are good ecological markers, as several species occur on most islands with each species occupying a specific or, at least, partially distinct habitat (e.g. Baptista et al., 1997; Gibbs et al., 2001; Wells & Wells, 2001). Several studies have described the past diversity and abundance of Columbidae in the Greater Antilles and Bahamian Archipelago. The initial work of Wetmore (1920, 1922a, 1922b, 1925, 1937) and more recent research in Cuba (Arredondo, 1984; Jiménez-Vázquez, 2001; Jiménez-Vázquez & Arrazcaeta, 2008, 2015; Olson & Wiley, 2016; Orihuela et al., 2020) and the Bahamas (e.g. Brodkorb, 1959; Olson & Hilgartner, 1982; Steadman & Franklin, 2020) has documented about 15 columbid taxa from late Pleistocene to historical period. However, in other areas of the Caribbean, namely the Lesser Antilles, the fossil and subfossil records remain poorly documented, comprising not more than 8 columbid taxa, most of which have been identified on the islands of Montserrat, Antigua and Sint Eustatius (Steadman et al., 1984a, 1984b; Van der Klift, 1992; Pregill et al., 1994). In other Lesser Antilles islands, columbid birds are usually not attributed beyond genus (Grouard, 2001, 2013; Newsom & Wing, 2004). In fact, identifying columbid bones from archaeological and paleontological sites to species remains difficult, especially for medium-sized species (Olson & Hilgartner, 1982; Steadman et al., 1984b; Goldberg, 1999; Olson, 2011). One reason for this is the substantial inter- and infraspecific variability that complicates reliably determining fossil remains, meaning that, while quantitatively well represented in fossil assemblages, a large number of columbid bones cannot be assigned to species. Moreover, only limited and partial data is available to identify the bones of several columbid species in the Caribbean and Bermuda (Wetmore, 1922a; Olson & Hilgartner, 1982; Steadman et al., 1984b; Worthy, 2001; Tellkamp, 2005; Olson, 2011).
OBJECTIVES
- Provide new osteological descriptions and quantitative data from the morphometric analysis of modern pigeons and doves for the reliable identification of Caribbean columbid bones from both archaeological and paleontological contexts.
MATERIALS AND METHODS
Specimens. The reference material for this study comes from the skeletal collection of the PACEA laboratory, University of Bordeaux, as well as the American Museum of Natural History of New York (AMNH), Musée Edgar Clerc (MEC), Muséum National d’Histoire Naturelle (MNHN), Natural History Museum (NHMUK), University of Florida-Florida Museum of Natural History (UF), and the Smithsonian Institution-National Museum of Natural History (USNM).
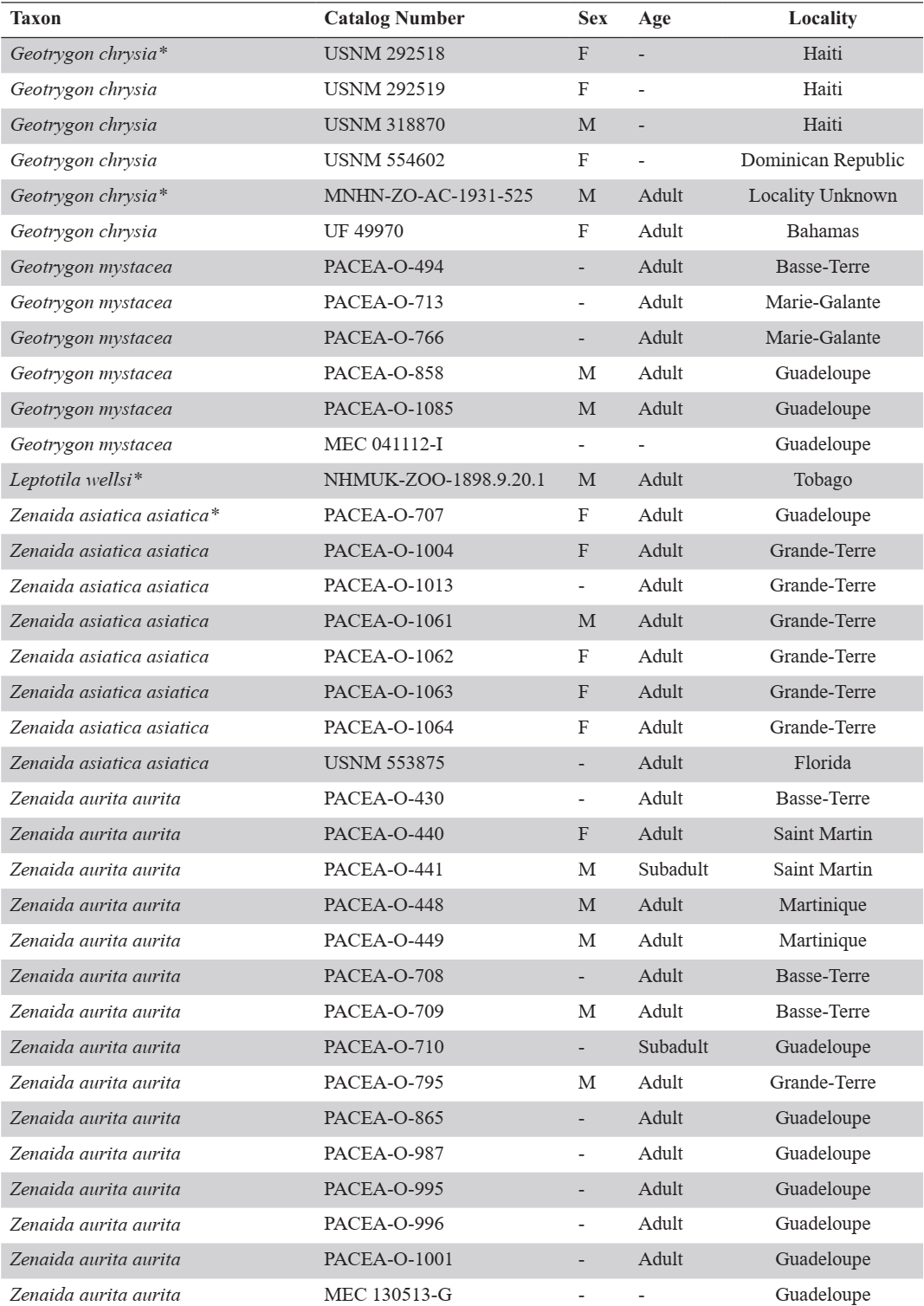
The sample included all pigeon species, resident or seasonal, currently present in the Antilles. The 3 species recently introduced to the Caribbean (Columba livia, Streptopelia decaocto and Streptopelia chinensis) and species with a limited distribution in the Western Caribbean (e.g., Columba caribaea, Geotrygon caniceps, Geotrygon versicolor) were excluded. The overall sample includes 80 more or less complete modern specimens of Columbidae (Appendix A), encompassing 5 genera, 12 species: the Scaly-naped Pigeon Patagioenas squamosa (n = 10), White-crowned Pigeon P. leucocephala (n = 11), Plain Pigeon P. inornata (n = 8), Common Ground Dove Columbina passerina (n = 2), Ruddy Quail-Dove Geotrygon montana (n = 5), Key West Quail-Dove G. chrysia (n = 7), Bridled Quail-Dove G. mystacea (n = 6), White-winged Dove Zenaida asiatica (n = 8), Zenaida Dove Z. aurita (n = 15), Eared Dove Z. auriculata (n = 5), and Mourning Dove Z. macroura (n = 2). A virtual specimen of the rare species Grenada Dove Leptotila wellsi was also included in the sample. Sex, age, and locality of origin were recorded for each specimen. Skeletal maturity is almost complete in some specimens, and several characters are evident even when the bird displayed juvenile plumage (subadult osteological specimens).
Methodology. We analysed 8 skeletal elements commonly found in the archaeological and paleontological record: coracoideum, scapula, humerus, ulna, carpometacarpus, femur, tibiotarsus and tarsometatarsus.
Bone orientation and descriptions of anatomical features follow the terminology of Baumel and Witmer (1993), with some exceptions. Osteological features on some elements of different European and Neotropical columbids identified by Wetmore (1922a), Fick (1974), Olson and Hilgartner (1982), Steadman et al. (1984b), Worthy (2001), Tellkamp (2005), and Tomek and Bochenski (2009) have been integrated into this study. The anatomical features used in the description of characters are illustrated for each bone, and characters are described respecting their anatomical position (from proximal to distal bone portions). The description and a photo are provided for the different states (from 2 to 4, A-D) of each morphological character. Specimens were assigned to intermediate states when they displayed less expressed characters or those falling between two states. The referred specimen, the percentage of states, and sample size (n) are provided for each taxon.
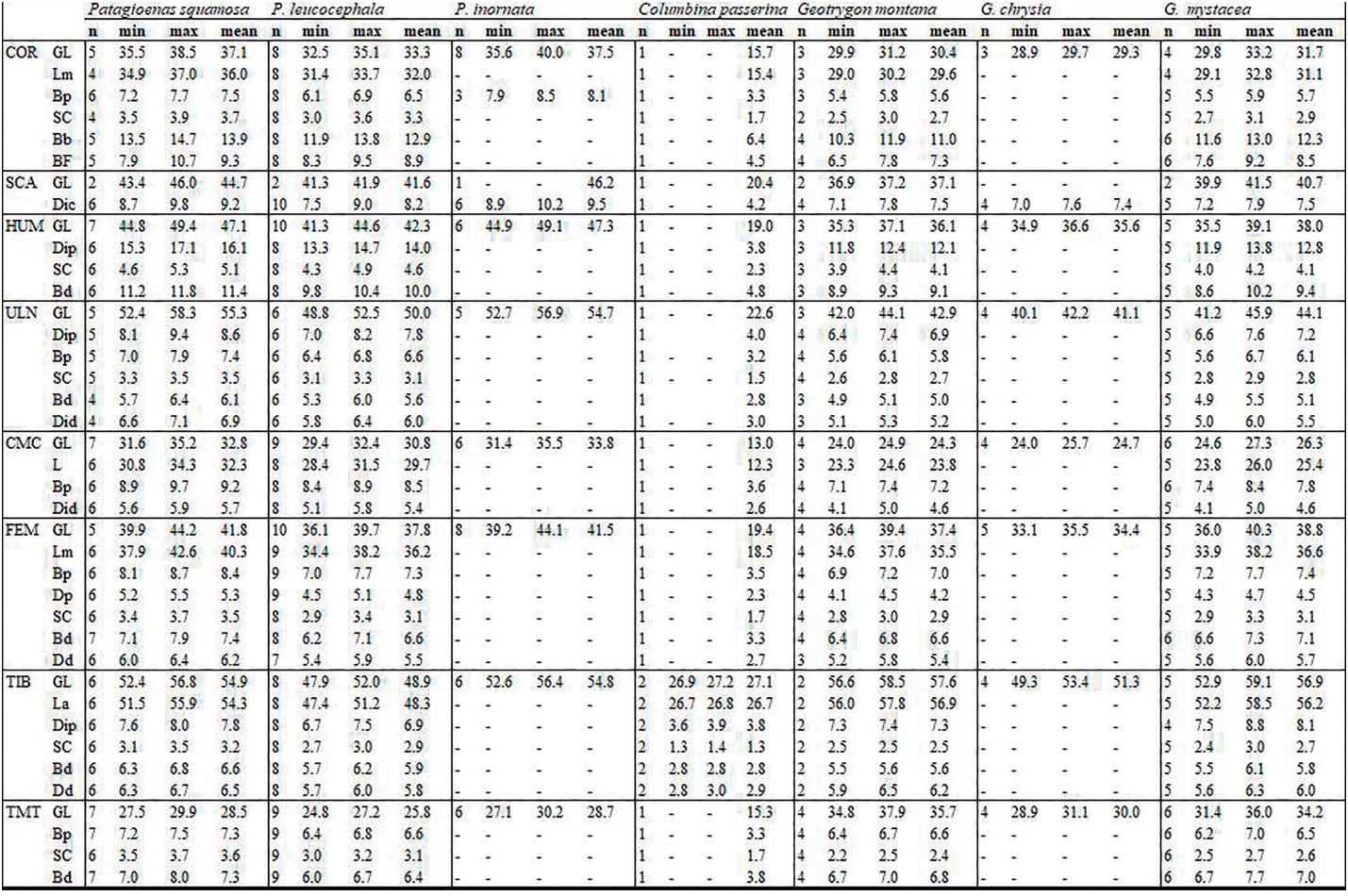
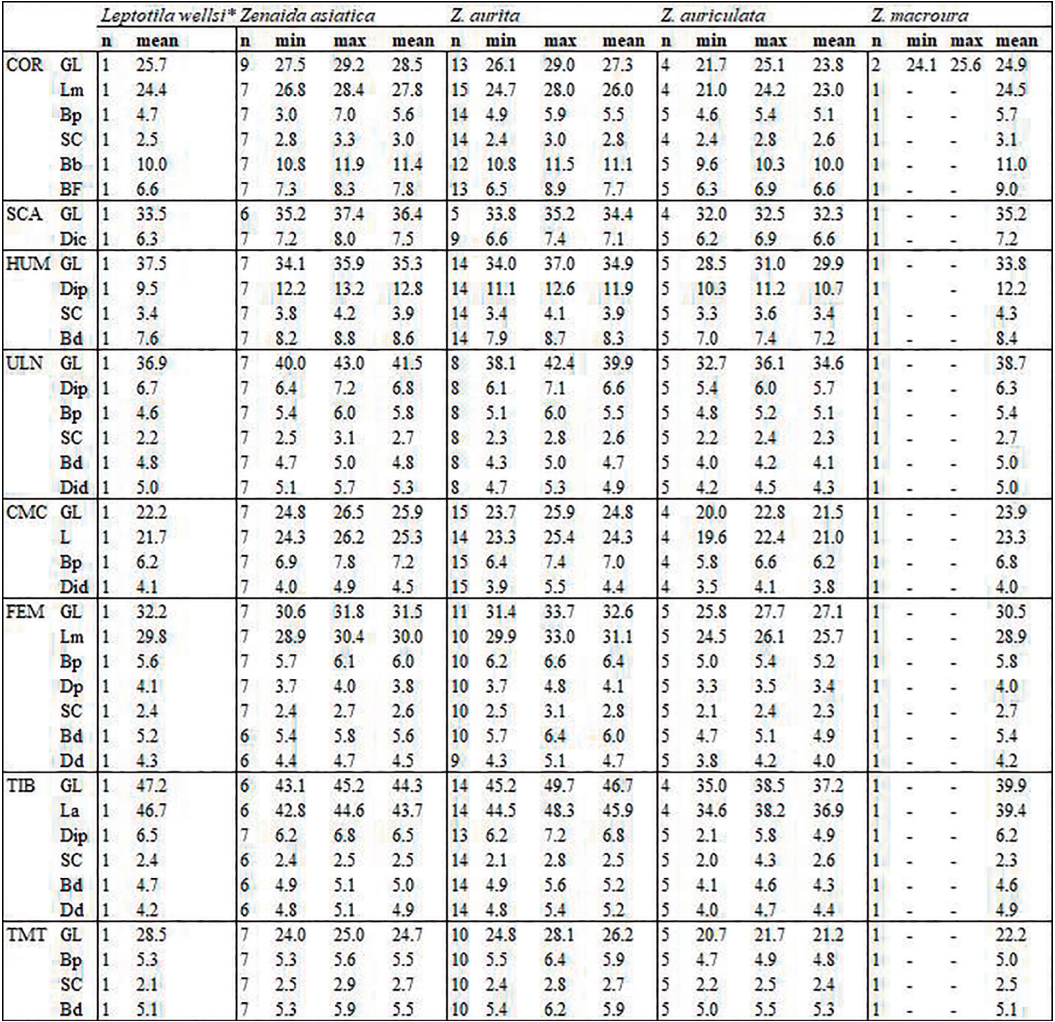
Measurements follow those described by von den Driesch (1976) but with two additions for the coracoideum: the proximal end (Bp) and smallest breadth (SC), (Appendix B). All bones were measured with a digital caliper to the nearest 0.1 mm and primarily concerned right-side bones. Mean, maximum, and minimal values of repeated measurements are provided in Appendix C. Some samples could not be fully measured due to damage (e.g. broken bone), anatomical connections, pathologies, or time contraints. For the 3-D models, the measurements were derived with the Avizo and Meshlab software packages. Measurements were compared with those provided by Goldberg (1999) for American Southwestern Columbidae (Z. asiatica, Z. macroura and C. passerina), with size classes based on those proposed by Steadman et al. (1984b) for pigeons and doves from the Lesser Antilles (P. squamosa, P. leucocephala, G. mystacea, G. montana, Z. aurita, C. passerina).
A single observer (MG) scored all criteria and measured all specimens. Only characters that occurred in at least 60 % of specimens are presented. Columbina passerina, the smallest species in the sample, cannot be confused with other species and is therefore included in the metric data excluded for describing specific characters. Detailed information (measurements and characters of each modern specimen) and illustrations of the 8 bones of the 4 genera are provided in the appendices (D-V).
RESULTS
A total of 42 morphological characters (7 for the coracoideum, 3 for the scapula, 6 for the humerus, 5 for the ulna, 11 for the carpometacarpus, 3 for the femur, 4 for the tibiotarsus, 3 for the tarsometatarsus) were selected. Due to the lack of sex data for numerous specimens, we excluded sexual dimorphism when discussing our results. However, male and female pigeons and doves are roughly similar in size, with appreciable dimorphism between sexes recorded only for Zenaida species (i.e. males larger than females; Wiley, 1991; Baptista et al., 1997).
Coracoideum
Coracoidea from 73 specimens were analyzed (Appendix D). When complete, this bone is easily distinguishable between genera. The coracoideum of Patagioenas has a much longer, broader, and robust shaft (corpus coracoidei) than the other 3 genera, whereas in Geotrygon, the shaft is long and narrow, and in Zenaida, it is short and wide (Fig. 1; Appendix E). Seven osteological characters and their states allow for a more detailed diagnosis (Fig 2). Five of these diagnostic characters concern the omal portion of the coracoideum, while the two others are typical of the distal portion and allow different Zenaida species to be distinguished (see also Appendix F). The percentage and distribution of states for each taxon are reported in Table I.
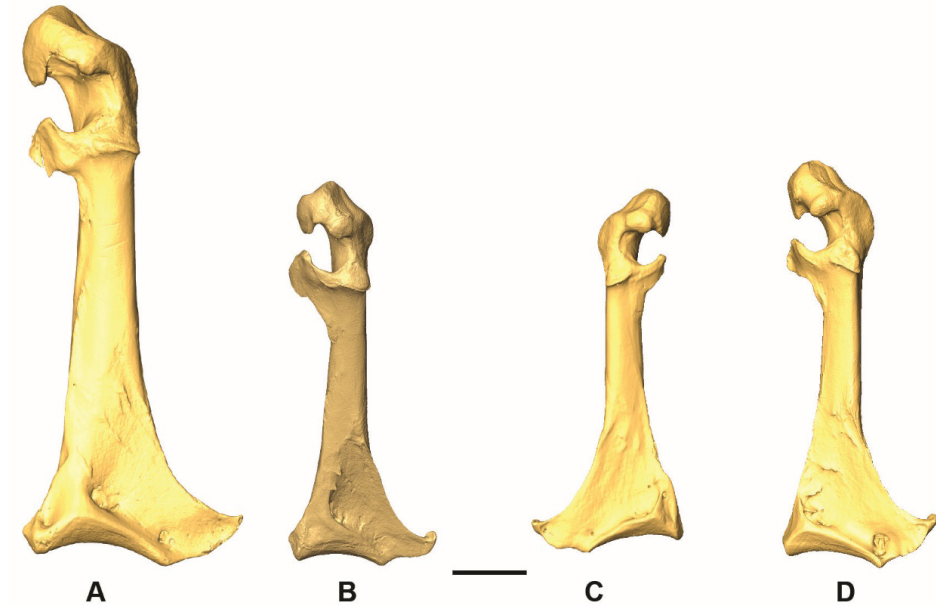
Margin of the facies articularis humeralis and cotyla scapularis (COR_FAH)
Referred material. Geotrygon mystacea PACEA-O-858 for state A and Zenaida aurita PACEA-O-865 for state B (Table I).
The margin of facies articularis humeralis and cotyla scapularis in dorsal view is broader in state A than in state B (Fig. 2).
All Zenaida specimens present a narrow margin, which sets them apart from the single L. wellsi specimen in the sample (Table I). Conversely, most Patagioenas and Geotrygon specimens exhibit a broad margin whose form differs according to species.
Shape of the area between the facies articularis clavicularis and the processus procoracoideus (COR_FAC)
Referred material. Patagioenas squamosa PACEA-O-442 for state A, Zenaida asiatica PACEA-O-1063 for state B, P. leucocephala PACEA-O-493 for state C, and Z. aurita PACEA-O-708 for state D (Table I).
In dorsal view, this element is oval in Geotrygon (Fig. 2; Table I). It is large in G. mystacea and most individuals of G. montana (state A) and narrow in G. chrysia (state D). Except for a single specimen of P. leucocephala, all Patagioenas specimens present a generally large, oval-shaped area between the articularis clavicularis and the processus procoracoideus.
In the single L. wellsi specimen and most Zenaida specimens, this area is rounded and narrow (state B), except in Z. asiatica, in which only two of the seven individuals present this character. In the other specimens assigned to this species it is narrow and oval, while in Z. auriculata it appears large and rounded (state C).
Pneumatic foramen (COR_PN)
Referred material. Patagioenas squamosa PACEA-O-442 for state A, Geotrygon montana PACEA-O-492 for state B, Zenaida auriculata PACEA-O-498 for state C, and Z. asiatica PACEA-O-707 for state D (Table I).
A pneumatic foramen is visible in medial view and can be found in the dorsal, central, or ventral portion of the bone. It is positioned dorsally (state A) in the upper part of the sulcus musculi supracoracoidei in Patagioenas (Fig. 2).
In Geotrygon, the presence of a pneumatic foramen is exclusive to G. montana, occurring centrally (state B) or in an intermediate position between the dorsal and midpoint of the bone (state A/B). This character is absent (state D) in the other two species (G. chrysia and G. mystacea) of this genus and in L. wellsi.
The absence of a marked pneumatic foramen is systematic in Zenaida, although a deep depression is occasionally present in the sulcus musculi supracoracoidei or below the facies articularis clavicularis (ventral position, state C): Z. aurita and Z. auriculata.
Tuberculum caudale (COR_TC)
Referred material. Zenaida asiatica PACEA-O-707 for state A, Patagioenas leucocephala PACEA-O-970 for state B, and Geotrygon montana PACEA-O-847 for state C (Table I).
In medial view, the tuberculum caudale (Fick, 1974) appears very prominent
(projects dorsally) in several cases, as in Zenaida asiatica (state A) and half of the coracoidea of Z. auriculata, whereas the two congeneric species have a prominent (state B) or non-prominent tuberculum (state C), (Fig. 2).
In Geotrygon, the tuberculum caudale is slightly or not prominent and absent in state A. While attributions to P. squamosa or P. leucocephala based on this character are less certain, the P. inornata primarily lacks a prominent tuberculum caudale. A single specimen of L. wellsi has a prominent tuberculum.
Processus acrocoracoideus (COR_PA)
Referred material. Geotrygon montana PACEA-O-847 for state A and Zenaida asiatica PACEA-O-707 for state B (Table I).
In ventral view, the processus acrocoracoideus in Geotrygon is more often distinctly raised (state A) and upwardly projected compared to state B (Fig. 2). This character is also observed in P. squamosa and some individuals of Z. aurita (Table I).
This element is less distinctly raised (state B) in P. inornata, Z. asiatica, Z. auriculata, and in the single Z. macroura and L. wellsi individuals. The processus acrocoracoideus is either distinctly raised or raised in P. leucocephala and Z. aurita.
Additionaly, 3 omal characters are described in the appendix (Appendix F).
Facies articularis sternalis near the angulus medialis (COR_FAS)
Referred material. Zenaida asiatica PACEA-O-707 for state A and Z. aurita PACEA-O-865 for state B (Table I).
This character and all of the following ones describe the area of the angulus medialis and allow different Zenaida species to be identified (Fig. 3).
In ventral view, the facies articularis sternalis near the angulus medialis is distinctively large for all individuals of Z. asiatica (state A, Fig. 3A), while this portion of the articular surface is quite narrow in Z. aurita (Fig. 3B), Z. auriculata (Fig. 3C), and Z. macroura (Fig. 3D).
Angulus medialis (COR_AM)
Referred material. Zenaida asiatica PACEA-O-707 for state A and Z. aurita PACEA-O-865 for state B (Table I).
In sternal view, the angulus medialis is straight in Z. asiatica (state A, Fig. 3E). All coracoidea of Z. aurita except one present a concavity on the labrum internum coracoidei (Livezey & Zusi, 2006), (state B, Fig. 3F). As all coracoidea present an intermediate state A/B (Fig. 3G-H), it is impossible to attribute specimens to either Z. auriculata or Z. macroura based on this character.
Coracoideum measurement
As expected, while genera are well differentiated, different species within each genus are less so (Fig. 4; Appendix C). Our results are in perfect agreement with those of Goldberg (1999), although the limited representation of certain taxa in our sample suggests a poor estimation of interspecific variability (Fig. 4). With that said, our data paints a more complex situation than previously assumed (Steadman et al., 1984b). Metrical data for the coracoideum does not separate Geotrygon mystacea and montana, and Geotrygon mystacea and Patagioenas leucocephala are only partially differentiable. In the latter case, despite a non-negligible overlap between the coracoideum measurements of these two taxa, the difference in mean coracoideum length is very close to being significant (t-student, p = 0.097). This is because smaller of the G. mystacea fall outside the size range of P. leucocephala while the larger of the latter taxon is outside the variability of the quail-doves. Overall, 5 distinct size classes are observable: P. squamosa = P. inornata > P. leucocephala ~= G. mystacea ~= G. montana > G. chrysia = Z. asiatica = Z. aurita > Z. auriculata = Z. macroura >> C. passerina.
Apart from being comparable in size to the different species of Zenaida, the small sample size for L. wellsi does not allow us to place this taxon within a particular size class.
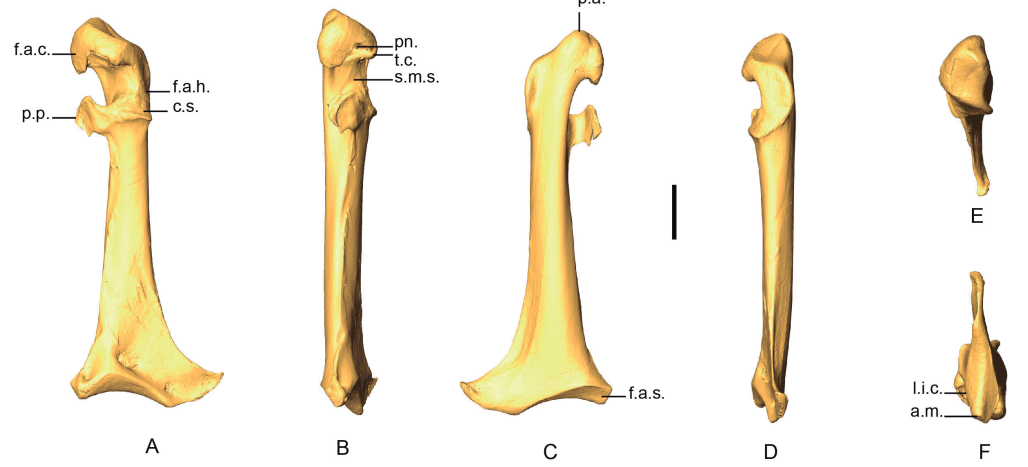
Figure 1. 3D - model of a right coracoideum of Patagioenas inornata USNM 226458 in dorsal (A), medial (B), ventral (C), lateral (D), proximal (E), and distal (F), views. Abbreviations: a.m., angulus medialis; c.s., cotyla scapularis; f.a.c., facies articularis clavicularis; f.a.h., facies articularis humeralis; f.a.s., facies articularis sternalis; l.i.c., labrum internum coracoidei; p.a., processus acrocoracoideus; pn., pneumatic foramen; p.p., processus procrocoracoideus; s.m.s., sulcus musculi supracoracoidei; t.c., tuberculum caudale. Scale bar equals 5 mm.

Figure 2. States of the osteological characters described for the coracoideum.
Table I. Coracoideum characters. Sample size (n) and percentage of states (A-D) and intermediate state (A/B, B/C, B/D) for each taxon
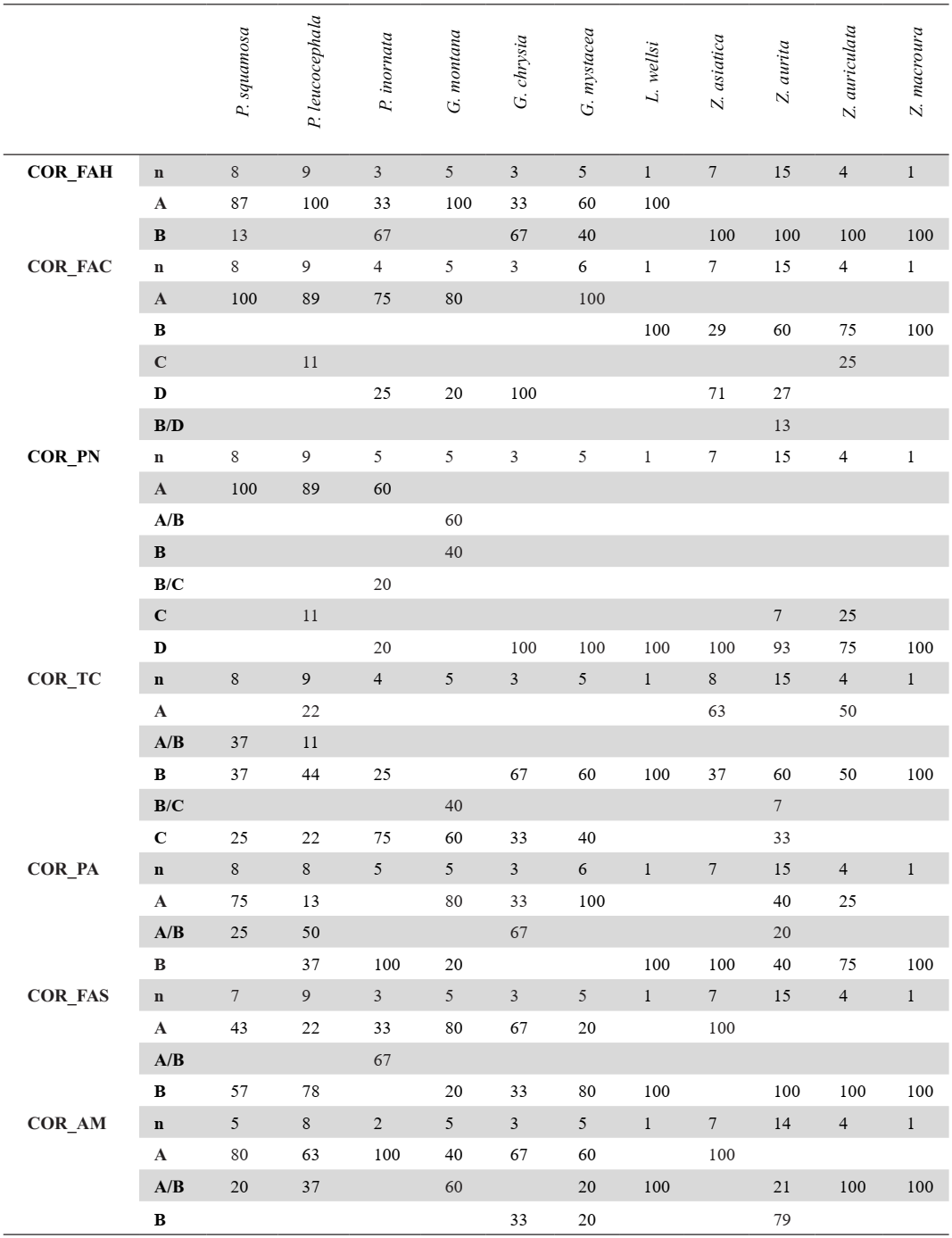
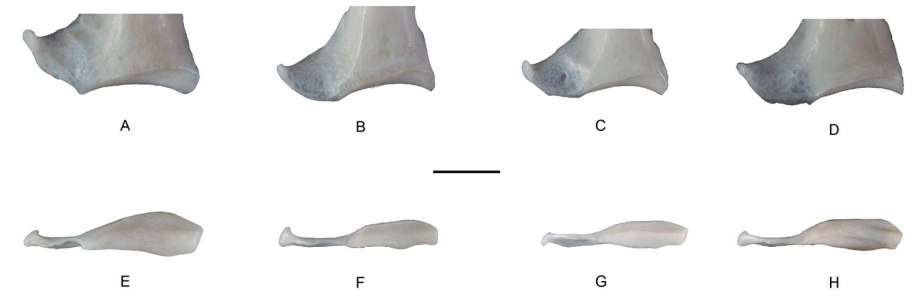
Figure 3. Distinctive characters of facies articularis sternalis and angulus-medialis in ventral (A-D) and sternal (E-H) views of coracoideum of Zenaida; A, E, Z. asiatica PACEA-O-707; B, F, Z. aurita PACEA-O-865; C, G, Z. auriculata PACEA-O-498; D, H, Z. macroura PACEA-O-493. Right side. Scale bar equals 1 cm.
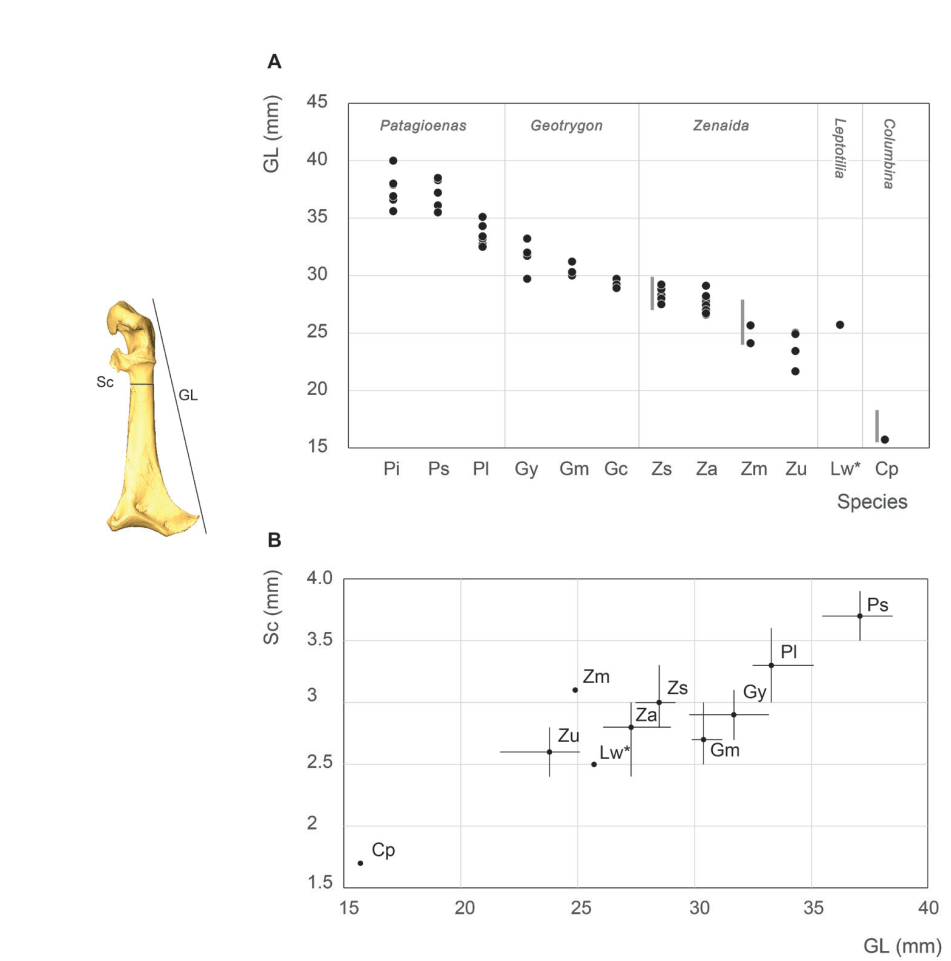
Figure 4. Coracoideum measurement in mm: GL, Greatest length (A), Sc, smallest breadth of the corpus and GL, Greatest length (B). See Appendix C. Abbreviations: Cp, Columbina passerina; Gc, Geotrygon chrysia; Gm, Geotrygon montana; Gy, Geotrygon mystacea; Lw, Leptotila wellsi; Pi, Patagioenas inornata; Pl, Patagioenas leucocephala; Ps, Patagioenas squamosa; Za, Zenaida aurita; Zm, Zenaida macroura; Zs, Zenaida asiatica; Zu, Zenaida auriculata. (*) 3D measurement; the grey vertical line in panel A indicates Goldberg’s measurements (1999).
Scapula
Scapulae from 66 specimens have been considered (Appendix G). The observations of complete bones demonstrate scapulae of Patagioenas (Fig. 5) and Zenaida to primarily exhibit a more curved ventral margin of the shaft (corpus scapulae) in medial or lateral view compared to those of the 2 other genera Geotrygon and Leptotila (Appendix H).
However, the substantial degree of inter-specific variability of this bone in the 4 genera complicates the recognition of diagnostic characters. Three osteological criteria and associated states of the articular portion of the scapula, two in articular view and one in medial view medial, are illustrated in Figure 6. The percentage and the distribution of states for each taxon are reported in Table II.
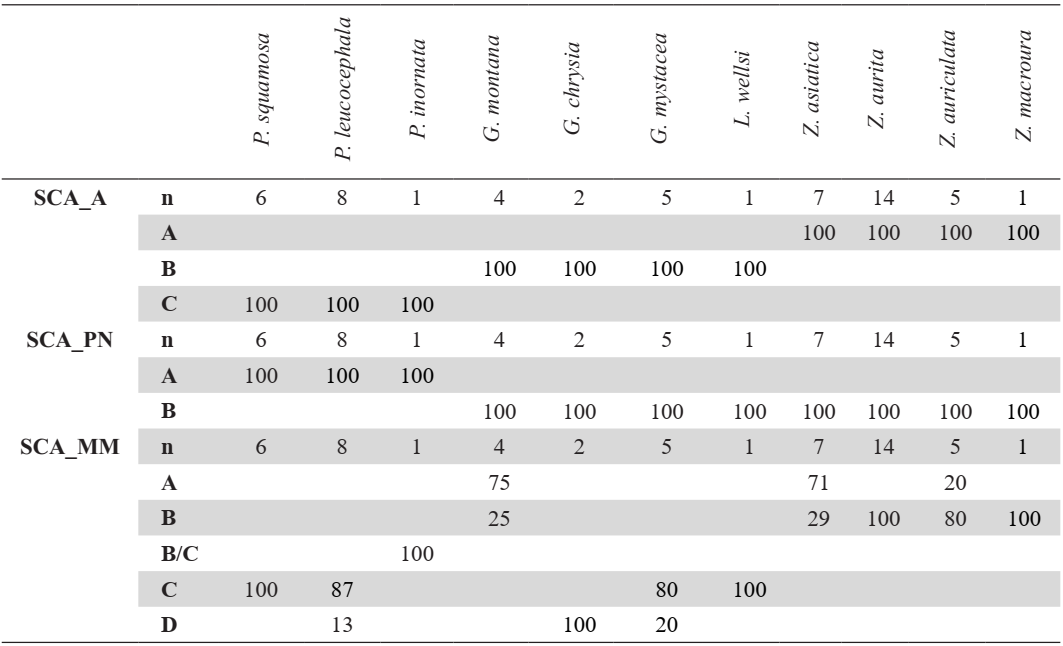
Shape of acromion (SCA_A)
Referred material. Zenaida aurita PACEA-O-708 for state A, Geotrygon montana PACEA-O-1055 for state B, and Patagioenas squamosa PACEA-O-442 for state C (Table II).
In medial view, the acromion is flat and rounded (state A) in Zenaida (Fig. 6). The medial apex is flattened, and the lateral apex is rounded while in state B the medial apex is flattened and the lateral apex is angular. This latter character is observed in Geotrygon and in L. wellsi (Appendix H).
In Patagioenas inornata and two congeneric species, the medial apex is larger than in other species and the lateral apex is rounded (state C, Fig. 6).
Pneumatic foramen (SCA_PN)
Referred material. Patagioenas squamosa PACEA-O-442 for state A and Z. aurita PACEA-O-708 for state B (Table II).
In articular view, a large pneumatic foramen is visible on the acromion (state A) in Patagioenas specimens, while in Geotrygon, Zenaida, and Leptotilia this foramen is absent (state B), (Fig. 6).
Medial margin between facies articularis humeralis and medial apex (SCA_MM)
Referred material. Geotrygon montana PACEA-O-847 for state A, Zenaida aurita PACEA-O-710 for state B, Patagioenas squamosa PACEA-O-442 for state C, and G. mystacea PACEA-O-766 for state D (Table II).
In articular view, the shape of the medial margin differs from one species to another (Fig. 6; Table II). This margin is convex and projects medially (state A) in 2 species: G. montana (except one scapula in Appendix I) and Z. asiatica.
In Z. aurita, Z. macroura, and most of the Z. auriculata individuals, the medial margin is concave (i.e. U-shaped) in state B. State C is slightly different but clearly distinguisable from state B; the two portions of the margin are more oblique and the margin is an open V-shape: P. squamosa, P. leucocephala, G. mystacea and L. wellsi.
One specimen of P. inornata exhibits characters of both B and C states. In rare cases, a relatively straight margin (state D) is present, as observed in the two G. chrysia specimens and the single specimens of P. leucocephala and G. mystacea.
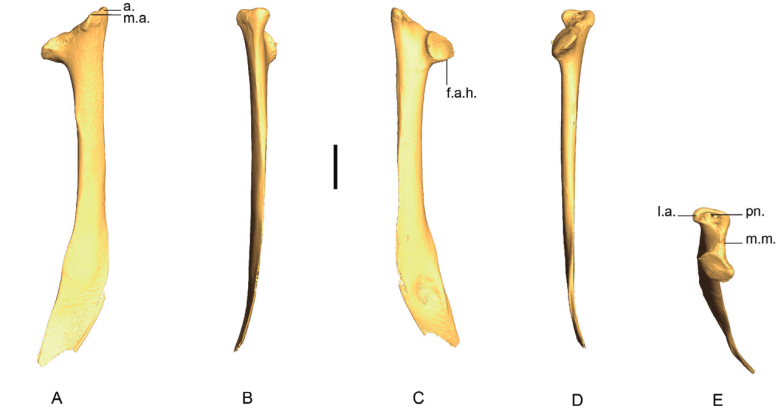
Figure 5. 3D - model of a right scapula of Patagioenas inornata USNM 290977 in medial (A), dorsal (B), lateral (C), ventral (D) and articular (E) views. Abbreviations: a., acromion; f.a.h., facies articularis humeralis; l.a., lateral apex; m.a., medial apex; m.m., medial margin; pn., pneumatic foramen. Scale bar equals 5 mm.
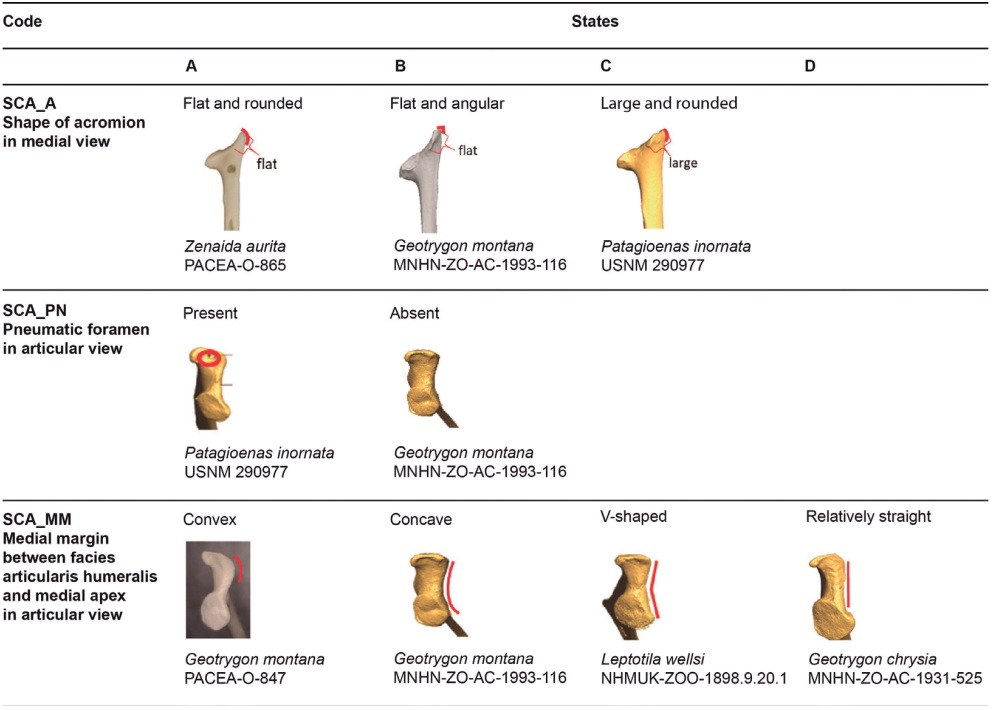
Figure 6. States of the osteological characters described for the scapula.
Table II. Scapula characters. Sample size (n) and percentage of states (A-D) and intermediate state (B/C) for each taxon
Scapula measurements
Given the fragility of its extremity, only a limited number of scapulae could be measured in their entirety (Appendix C). Despite this limitation, the proximal width of the scapula is greater in Patagioena compared to the other three genera (Fig. 7).
Scapula measurements were similar to those reported by Goldberg (1999) but differ from those of Steadman et al. (1984b); the larger P. leucocephala has a longer scapula than G. mystacea, while scapula length in G. mystacea and G. montana appear indistinguishable and
Z. aurita has a shorter scapula compared to G. montana.
The same five size classes identified for the coracoideum are equally evident in the greatest length of scapula, including the same degree of uncertainty for the position of L. wellsi which, reflects the single scapula measurement for this species.
Humerus
Humeri from 66 specimens have been considered (Appendix I). Extremely difficult to distinguish between different genera, even when complete, the humerus of Patagioenas is generally long and robust (Fig. 8). Distinguishing the genera Geotrygon and Zenaida based on metrical data is difficult, even if the humerus of Geotrygon mystacea is generally longer. The humerus is more curved in Zenaida compared to the other genera (Appendices J-K).
Six osteological criteria are defined and illustrated (Fig. 9). The distribution of the different states by species is reported in Table III. Four of these diagnostic characters were found in the proximal portion of the bone, while the two others describe the distal portion.
Caput humeri (HUM_CH)
Referred material. Patagioenas squamosa PACEA-O-442 for state A and Geotrygon montana MNHN-ZO-AC-1993-116 for state B (Table III).
In cranial view, the caput humeri projects ventrally beyond the shaft axis (state A) in Patagioenas as well as in G. chrysia (Fig. 9). The caput is more centrally positioned (state B) in G. montana, G. mystacea, and L. wellsi. This character is present in the two states in Zenaida specimens.
Ventral margin of the shaft and crista bicipitalis (HUM_VM)
Referred material. Patagioenas squamosa PACEA-O-442 for state A, P. inornata USNM 290977 for state B, and Zenaida asiatica PACEA-O-707 for state C (Table III).
In cranial view, the crista bicipitalis and the corpus humeri form a large, rounded arch (state A) in Patagioenas and Geotrygon (Fig. 9). In state B, the ventral margin is more tightly rounded than in state A (L. wellsi). In Zenaida, the ventral margin is angular (state C, see appendix K for an additional character of this genus).
Tuberculum dorsale (HUM_TD)
Referred material. Patagioenas squamosa PACEA-O-486 for state A, Zenaida aurita PACEA-O-710 for state B, P. leucocephala PACEA-O-822 for state C, and G. mystacea PACEA-O-494 for state D (Table III).
This character was recognized by Steadman et al. (1984b) as distinguishing P. leucocephala from G. mystacea. Our results are consistent with this distinction and also allow Zenaida to be individualized from Patagioenas.
The tuberculum dorsale can be wide and well developed (state A) or narrow (state B) and in many cases is associated with a depression in the cranial margin of the crista deltopectoralis (states C-D), (Fig. 9). Several obvious differences are evident on the two reference specimens of Patagioenas and Zenaida, although numerous variations between the two genera make it difficult to refer some specimens to a state. Moreover, in Geotrygon, a well-developed depression is evident in G. mystacea but is less marked in G. montana, where a slight depression is present. Therefore, some specimens of this genus present intermediate states (state B/D).
However, the tuberculum dorsale is generally well developed in Patagioenas (state A) and in some cases projects cranially and dorsally. In Zenaida, this element is narrow (states B or D) and in L. wellsi it is narrow with a depression (state D).
Ventral margin of the sulcus ligamentus transversus of the caput humeri (HUM_SLT)
Referred material. Geotrygon mystacea PACEA-O-494 for state A, G. montana MNHN-ZO- AC-1993-116 for state B and Zenaida aurita PACEA-O-710 for state C (Table III).
This character is visible in both proximal and ventral view and allows G. mystacea to be distinguished from G. montana. In the first species, the ventral margin of the sulcus ligamentus transversus of the caput humeri has a rounded profile (state A) while in G. montana this margin is oblique (state B, Fig. 9). A straighter margin (state C) is evident in some specimens of Zenaida.
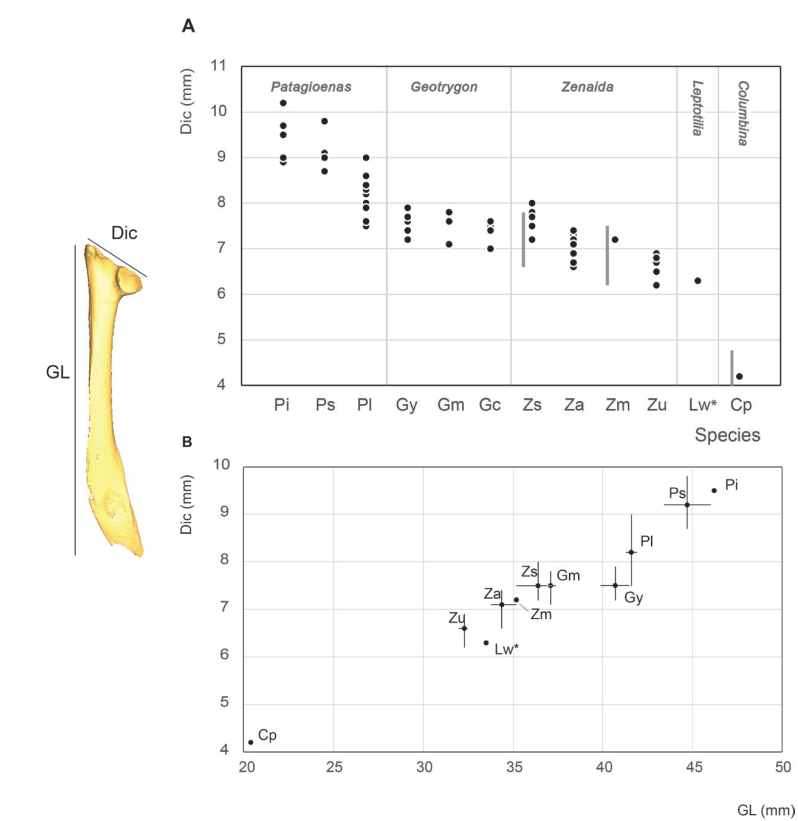
Figure 7. Scapula measurement in mm: Dic, (Greatest) cranial diagonal (A), Dic, (Greatest) cranial diagonal and GL, Greatest length (B). See Appendix C. Abbreviations: Cp, Columbina passerina; Gc, Geotrygon chrysia; Gm, Geotrygon montana; Gy, Geotrygon mystacea; Lw, Leptotila wellsi; Pi, Patagioenas inornata; Pl, Patagioenas leucocephala; Ps, Patagioenas squamosa; Za, Zenaida aurita; Zm, Zenaida macroura; Zs, Zenaida asiatica; Zu, Zenaida auriculata. (*) 3D measurement; the grey vertical line in panel A indicates Goldberg’s measurements (1999).
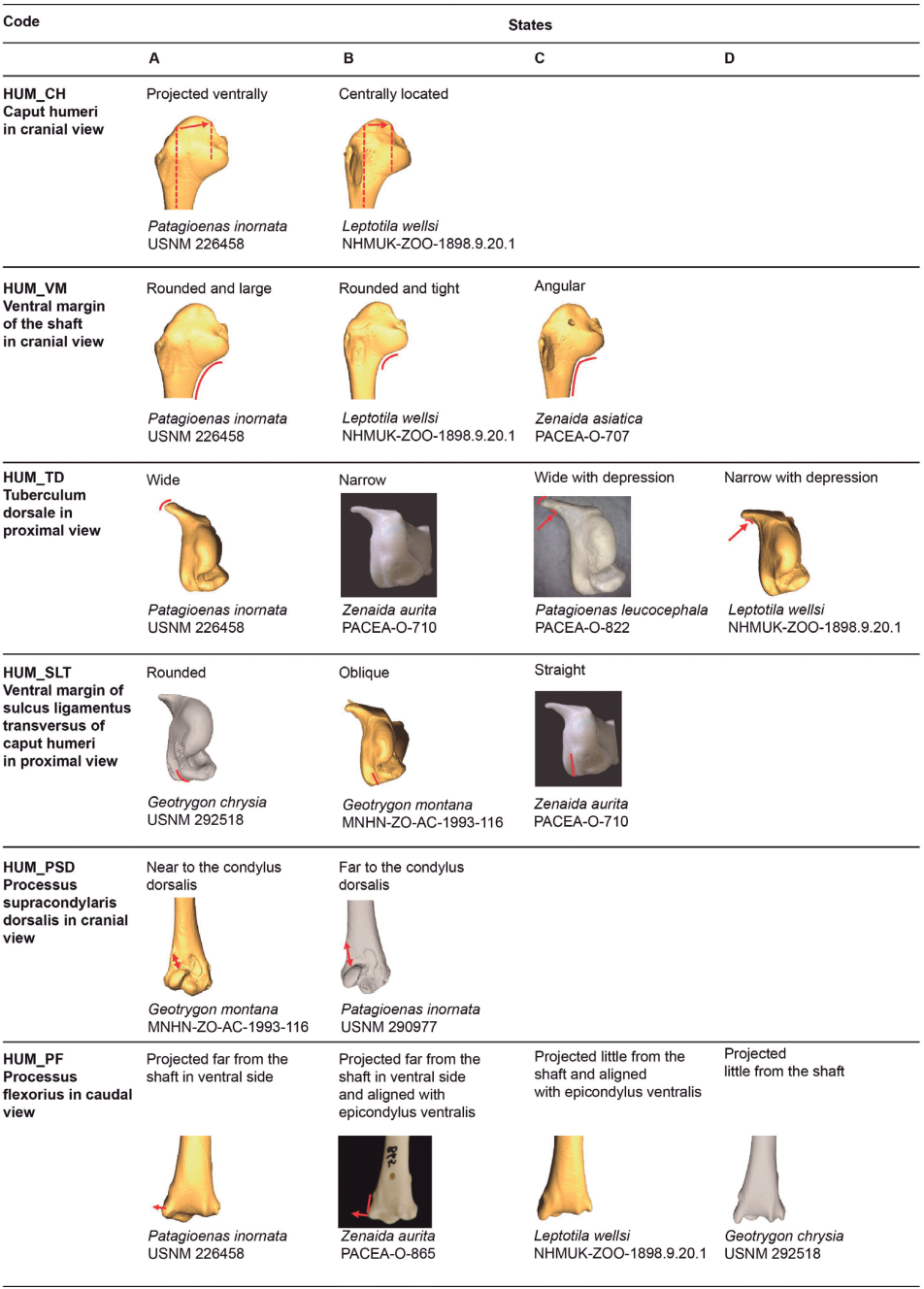
Figure 8. 3D - model of a right humerus of Patagioenas inornata USNM 226458 in cranial (A), dorsal (B), caudal (C), ventral (D), proximal (E), and distal (F), views. Abbreviations: c.b. crista bicipitalis; c.d.p., crista deltopectoralis; c.h., caput humeri; co.d., condylus dorsalis; e.v., epicondylus ventralis; p.f., processus flexorius; p.s.d., processus supracondylaris dorsalis; s.l.t., sulcus ligamentus transversus of caput humeri; t.d., tuberculum dorsale; v.m., ventral margin. Scale bar equals 1 cm.
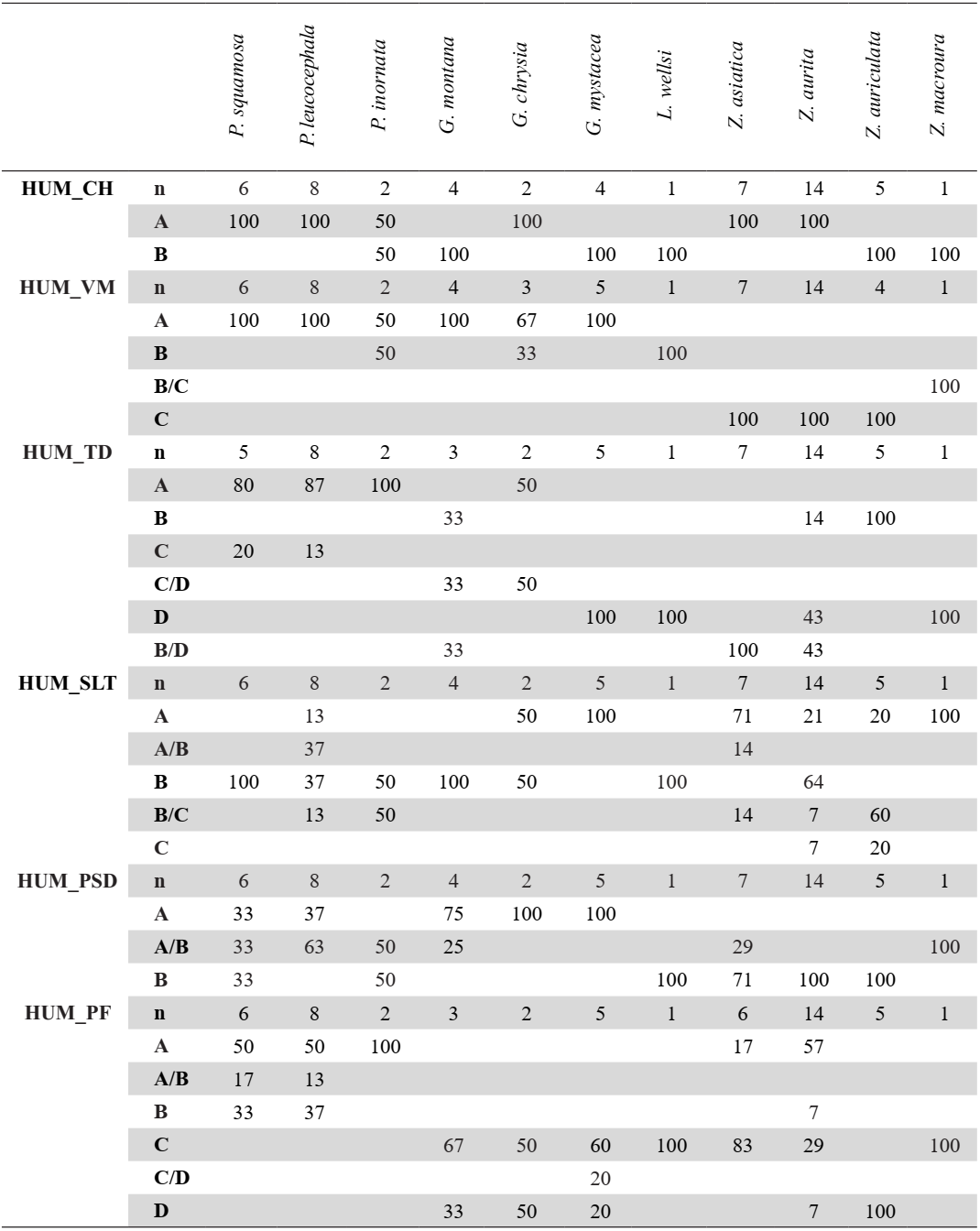
Figure 9. States of the osteological characters described for the humerus.
Table III. Humerus characters. Sample size (n) and percentage of states (A-D) and intermediate states (A/B, B/C, C/D, B/D) for each taxon
Processus supracondylaris dorsalis (HUM_PSD)
Referred material. Geotrygon montana MNHN-ZO-AC-1993-116 for state A and Zenaida asiatica PACEA-O-707 for state B (Table III).
The processus supracondylaris dorsalis (ectepicondylar prominence) has been previously used by Olson & Hilgartner (1982) to distinguish G. chrysia from Z. aurita. We equally observed this character in other species, indicating it to reliably distinguish the two genera Geotrygon and Zenaida. As in G. chrysia, the processus supracondylaris dorsalis is closer to the condylus dorsalis than in the two other Geotrygon species (state A, Fig. 9). In Zenaida, the processus supracondylaris dorsalis is positioned away from the condylus dorsalis (state B).
Processus flexorius (HUM_PF)
Referred material. Patagioenas squamosa PACEA-O-442 for state A, Zenaida aurita PACEA-O-865 for state B, Geotrygon mystacea PACEA-O-494 for state C, and G. montana PACEA-O-847 for state D (Table III).
The processus flexorius projects significantly from the shaft on the ventral side in Patagioenas (state A). In some cases, it is aligned with the epicondylus ventralis (state B) (Fig. 9). Conversely, in Geotrygon and L. wellsi, the processus flexorius projects only slightly from the shaft and is either aligned (state C) or unaligned (state D) with the epicondylus ventralis. This character is present in the four states in Zenaida specimens. Additionaly, one distal character is described in the appendix (Appendix K).
Humerus measurement
Our humerus measurements for Z. asiatica do not differ from those of Goldberg (1999) (Fig. 10). On the other hand, our Z. macroura and C. passerina specimens fall outside the variability documented for these two species in the southwestern United States in that they are, respectively, slightly larger and smaller than the specimens reported by Goldberg (1999).
Humerus length measurements are more evenly distributed compared to the two bones of the upper girdle (coracoideum and scapula). The different size classes are clearly evident in all three bones, although some differences in the distributions of taxa within each category are evident; humerus size in P. squamosa and P. inornata fall within the variability of large pigeons while P. leucocephala is within the small pigeon category. All Geotrygon specimens, the large Zenaida (Z. asiatica and Z. aurita) and Leptotila are classed as large doves. The small doves, Z. macroua, Z. auriculata and C. passerina, are easily distinguished by the very small size of their humerus. Unlike Steadman et al. (1984b) data, our measurements do not distinguish G. mystacea, G. montana and Z. aurita.
Ulna
Ulnae from 63 specimens have been considered (Appendix L). The complete ulnae of Patagioenas are longer and their shafts are more robust than the other 3 genera (Fig. 11; Appendix M). Distinguishing Geotrygon from Zenaida based on size is, however, difficult, even if the Geotrygon specimens are generally longer. Several morphological differences do exist between these two genera. We described 5 osteological criteria (4 on the proximal and 1 on the distal parts) for the 4 genera (Fig. 12) that distinguish different Geotrygon species, but which do not always separate them from certain forms of Zenaida. It is equally difficult to consistently distinguish different Zenaida species due to the variable expression of these characters (Table IV).
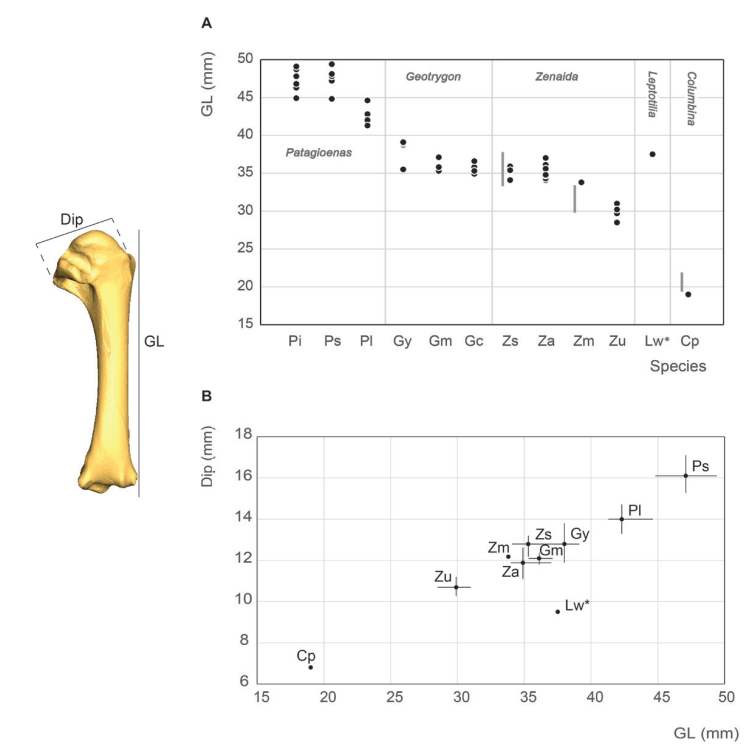
Figure 10. Humerus measurement in mm: GL, Greatest length (A), Dip, (Greatest) diagonal of the proximal end and GL, Greatest length (B). See Appendix C. Abbreviations: Cp, Columbina passerina; Gc, Geotrygon chrysia; Gm, Geotrygon montana; Gy, Geotrygon mystacea; Lw, Leptotila wellsi; Pi, Patagioenas inornata; Pl, Patagioenas leucocephala; Ps, Patagioenas squamosa; Za, Zenaida aurita; Zm, Zenaida macroura; Zs, Zenaida asiatica; Zu, Zenaida auriculata. (*) 3D measurement; the grey vertical line in panel A indicates Goldberg’s measurements (1999).
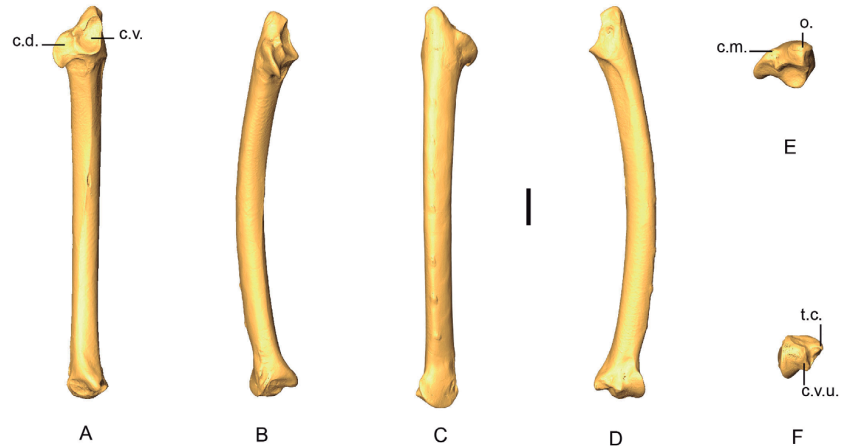
Figure 11. 3D - model of a right ulna of Patagioenas inornata USNM 226458 in cranial (A), dorsal (B), caudal (C), ventral (D), proximal (E), and distal (F), views. Abbreviations: c.m., caudal margin; c.d., cotyla dorsalis; c.v., cotyla ventralis; c.v.u., condylus ventralis ulnae; o., olecranon; t.c., tuberculum carpale. Scale bar equals 5 mm.
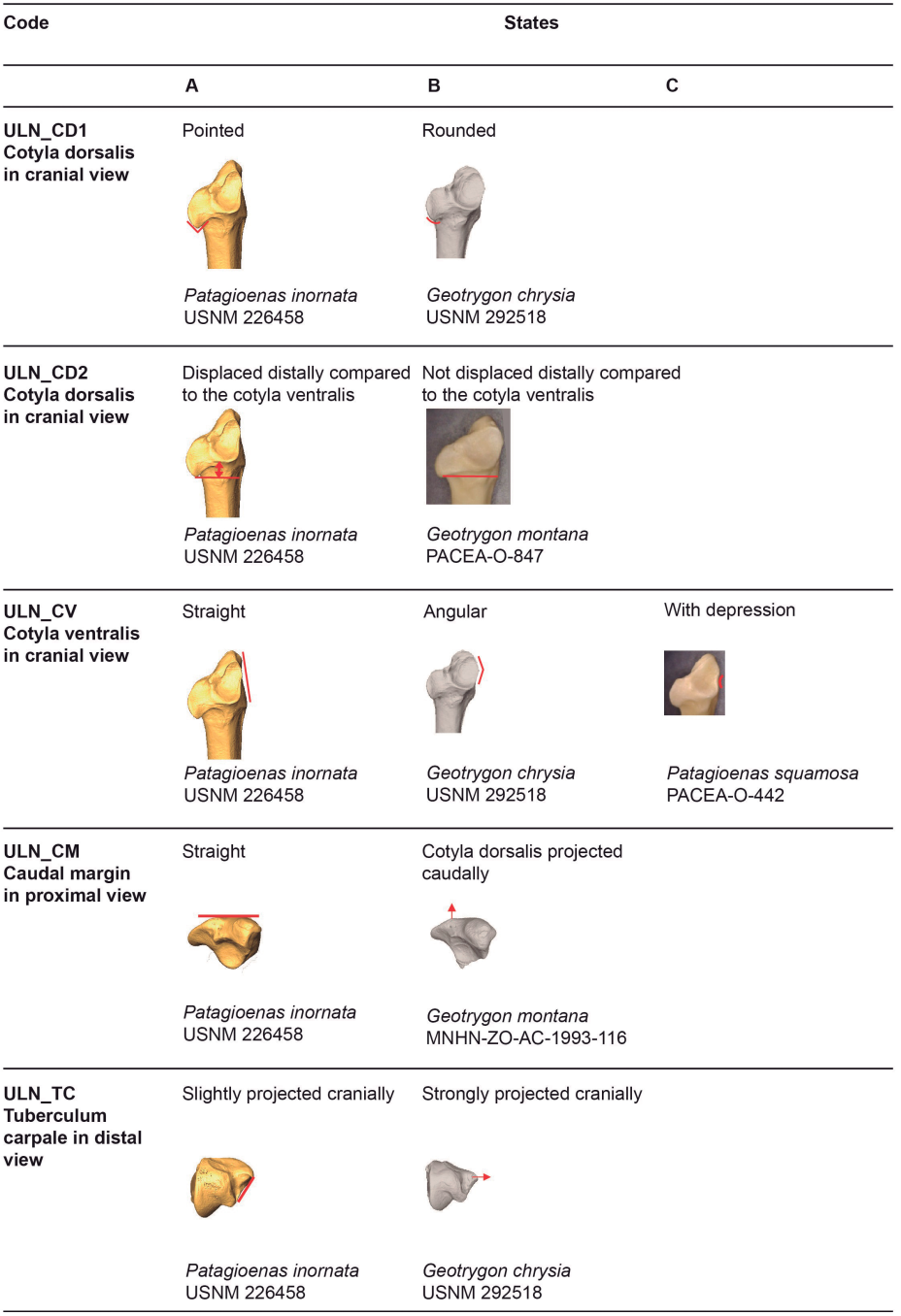
Figure 12. States of the osteological characters described for the ulna.
Table IV. Ulna characters. Sample size (n) and percentage of states (A-D) and intermediate states (A/B, B/C) for each taxon

Cotyla dorsalis (ULN_CD1)
Referred material. Patagioenas leucocephala PACEA-O-493 for state A and Geotrygon montana PACEA-O-847 for state B (Table IV).
The distal part of the cotyla dorsalis, in cranial view, has a pointed margin on most columbid specimens in our sample (state A), (Fig. 12). Some specimens exhibit a rounded cotyla dorsalis (state B): all specimens of G. montana, some of the P. squamosa and P. inornata specimens, as well as G. chrysia, L. wellsi and Z. macroura.
Cotyla dorsalis (ULN_CD2)
Referred material. Geotrygon mystacea PACEA-O-494 for state A and G. montana PACEA-O-847 for state B (Table IV).
In cranial view, the cotyla dorsalis is displaced distally compared to the cotyla ventralis (state A) or positioned at the same level (state B); (Fig. 12). In Geotrygon, this character is distinctive of G. montana.
Cotyla ventralis (ULN_CV)
Referred material. Patagioenas leucocephala PACEA-O-821 for state A, Zenaida aurita PACEA-O-865 for state B, and P. squamosa PACEA-O-442 for state C (Table IV).
In cranial view, the ventral margin of the cotyla ventralis is relatively straight and vertical (state A) in the L. wellsi specimen as well as several other species (Fig. 12). This state is absent from Geotrygon and rare in the other genera. In Geotrygon, the ventral margin is angular, the distal part of the ventral margin is relatively straight, and the proximal portion near the olecranon is oblique (state B). In addition, the cotyla ventralis of G. montana is more curved than in G. mystacea.
The cotyla ventralis of most P. squamosa specimens projects ventrally and has a depression in the distal part (state C). Three P. leucocephala specimens and four Zenaida specimens also exhibit this character.
Caudal margin of proximal end (ULN_CM)
Referred material. Patagioenas squamosa PACEA-O-442 for state A and Zenaida aurita PACEA-O-865 for state B (Table IV).
In the proximal view, the caudal margin of the proximal portion of the ulna is relatively straight and the cotyla dorsalis is aligned with the olecranon (state A) or projects caudally (state B), (Fig. 12). This latter state is observed in most Geotrygon specimens except for G. chrysia, in Zenaida apart from Z. auriculata and in P. squamosa.
Problematic species: no characteristic states in P. leucocephala.
Tuberculum carpale (ULN_TC)
Referred material. Patagioenas squamosa PACEA-O-442 for state A and Geotrygon montana PACEA-O-492 for state B (Table IV).
In distal view, the tuberculum carpale is slightly projected cranially and the cranial margin between the condylus ventralis ulnae and the tuberculum carpale is relatively straight (state A), (Fig. 12). This state is observed in most of the Patagioenas and Zenaida specimens, and in L. wellsi. The tuberculum projects heavily cranially and the cranial margin is angular in Geotrygon (state B).
Ulna measurement
Our ulna and humerus measurements differ in the same way as those reported by Goldberg (1999), namely a smaller-winged C. passerina specimen and a larger-winged Z. macroura specimen (Fig. 13).
The same five groupings of taxa for the humerus sample are equally evident amongst the ulna although slightly less clearly expressed; the largest small pigeons (P. leucocephala) are indistinct from the smallest individuals of the large pigeons (P. squamosa and P. inornata). Moreover, L. wellsi is closer to the small rather than the large doves; however, it should be noted that is this based on a single specimen of L. wellsi in our sample
Like the humerus, G. mystacea, G. Montana, and Z. aurita cannot be distinguished based on ulna length, an observation inconsistent with the data reported by Steadman et al. (1984b).
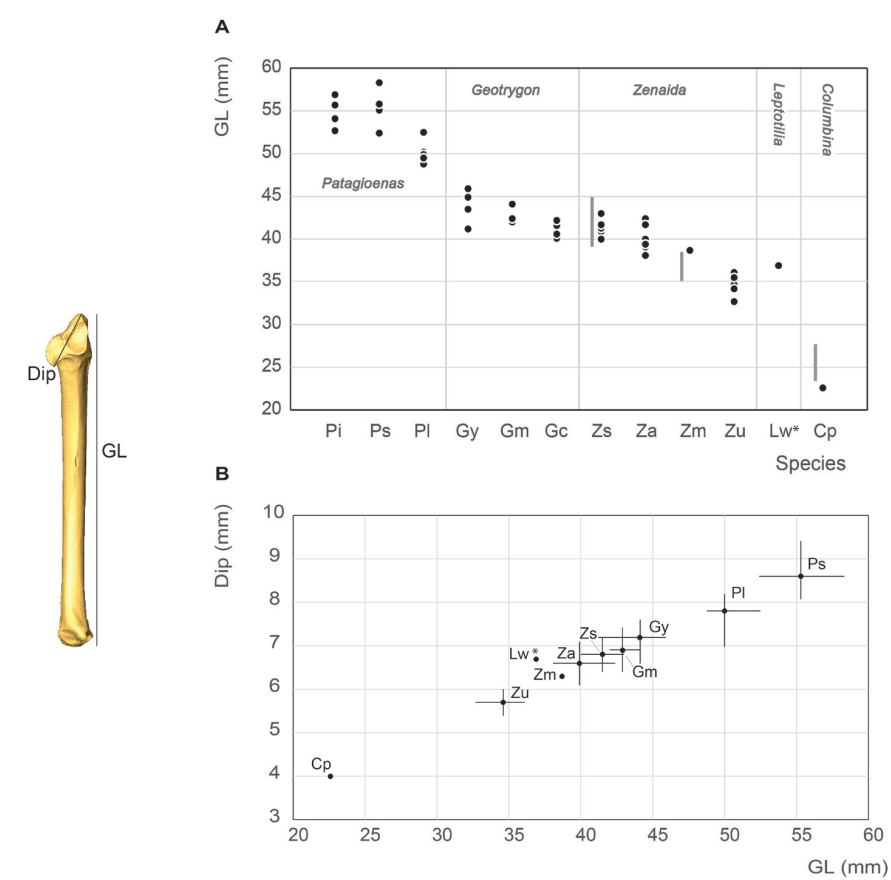
Figure 13. Ulna measurement in mm: GL, Greatest length (A), Dip, diagonal of the proximal end and GL, Greatest length (B). See Appendix C. Abbreviations: Cp, Columbina passerina; Gc, Geotrygon chrysia; Gm, Geotrygon montana; Gy, Geotrygon mystacea; Lw, Leptotila wellsi; Pi, Patagioenas inornata; Pl, Patagioenas leucocephala; Ps, Patagioenas squamosa; Za, Zenaida aurita; Zm, Zenaida macroura; Zs, Zenaida asiatica; Zu, Zenaida auriculata. (*) 3D measurement; the grey vertical line in panel A indicates Goldberg’s measurements (1999).
Carpometacarpus
Our sample comprises 68 specimens (Appendix N). The complete carpometacarpi of Patagioenas are longer and more robust than those of other genera (Fig. 14; Appendix O). Morphological differences between the 4 genera are based on 11 osteological criteria, which also are used to identify species (Fig. 15; Table V). Seven characters describe the proximal part of the bone, 1 the shaft and 3 the distal portion.
Dorsal margin of the os metacarpale minus near trochlea carpalis (CMC_OMM)
Referred material. Geotrygon mystacea PACEA-O-494 for state A and Patagioenas squamosa PACEA-O-442 for state B (Table V).
In dorsal view, the margin of the os metacarpale minus in its proximal part, near the trochlea carpalis, is wide (state A) in L. wellsi, G. chrysia, G. mystacea and some G. montana (Fig. 15, State 1). It is narrower (state B or intermediate state A/B) in the other specimens or species studied.
Trochlea carpalis (CMC_TC1)
Referred material. Patagioenas leucocephala PACEA-O-970 for state A, Geotrygon mystacea PACEA-O-494 for state B, G. montana PACEA-O-492 for state C, and Zenaida auriculata PACEA-O-498 for state D (Table V).
On the proximal portion of carpometacarpus, the trochlea carpalis in dorsal view is well-developed upwards (projecting well beyond the processus extensorius) and rounded (state A) in P. inornata (Fig. 15, State 1). It is developed upwards (projecting slightly from the processus extensorius) and flattened (state B) in G. mystacea and part of the P. leucocephala specimens. Some specimens of Eastern Caribbean columbids have a weakly developed trochlea (reaching approximately the same level as the processus extensorius) that is either rounded, as in some Patagioenas, Geotrygon and Zenaida species (state C), or flattened, as in Leptotila wellsi (state D).
Problematic species: no characteristic states in Z. aurita.
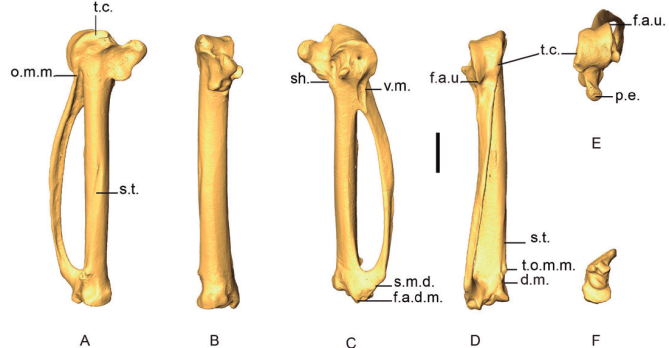
Figure 14. 3D - model of a right carpometacarpus of Patagioenas inornata USNM 226458 in dorsal (A), caudal (B), ventral (C), cranial (D), proximal (E), and distal (F), views. Abbreviations: d.m., dorsal margin; f.a.d.m., facies articularis digitalis major; f.a.u, facies articularis ulnocarpalis; o.m.m., os metacarpale minus; p.e., processus extensorius; sh., shaft; s.m.d., symphysis metacarpalis distalis; s.t., sulcus tendinous; t.c, trochlea carpalis; t.o.m.m., tuberosity of os metacarpale majus; v.m., ventral margin. Scale bar equals 5 mm.
Distal dorsal margin of trochlea carpalis (CMC_TC2)
Referred material. Patagioenas squamosa PACEA-O-442 for state A, Zenaida a. asiatica PACEA-O-707 for state B and Geotrygon montana PACEA-O-492 (Table V).
In caudal view, there is a distinct eminence on the distal dorsal margin of the trochlea carpalis (state A) in all or most of the observed specimens of P. inornata, G. chrysia and G. mystacea (Fig. 15, State 1). This eminence is less prominent (state B) in Z. asiatica, Z. aurita and Z. macroura. The dorsal margin does not present an eminence (state C) in
G. montana and L. wellsi.
Problematic species: depending on the individual, the 3 states are observed in P. leucocephala, and this eminence is relatively indistinct or absent in Z. auriculata. P. squamosa presents a distinct eminence or no eminence at all.
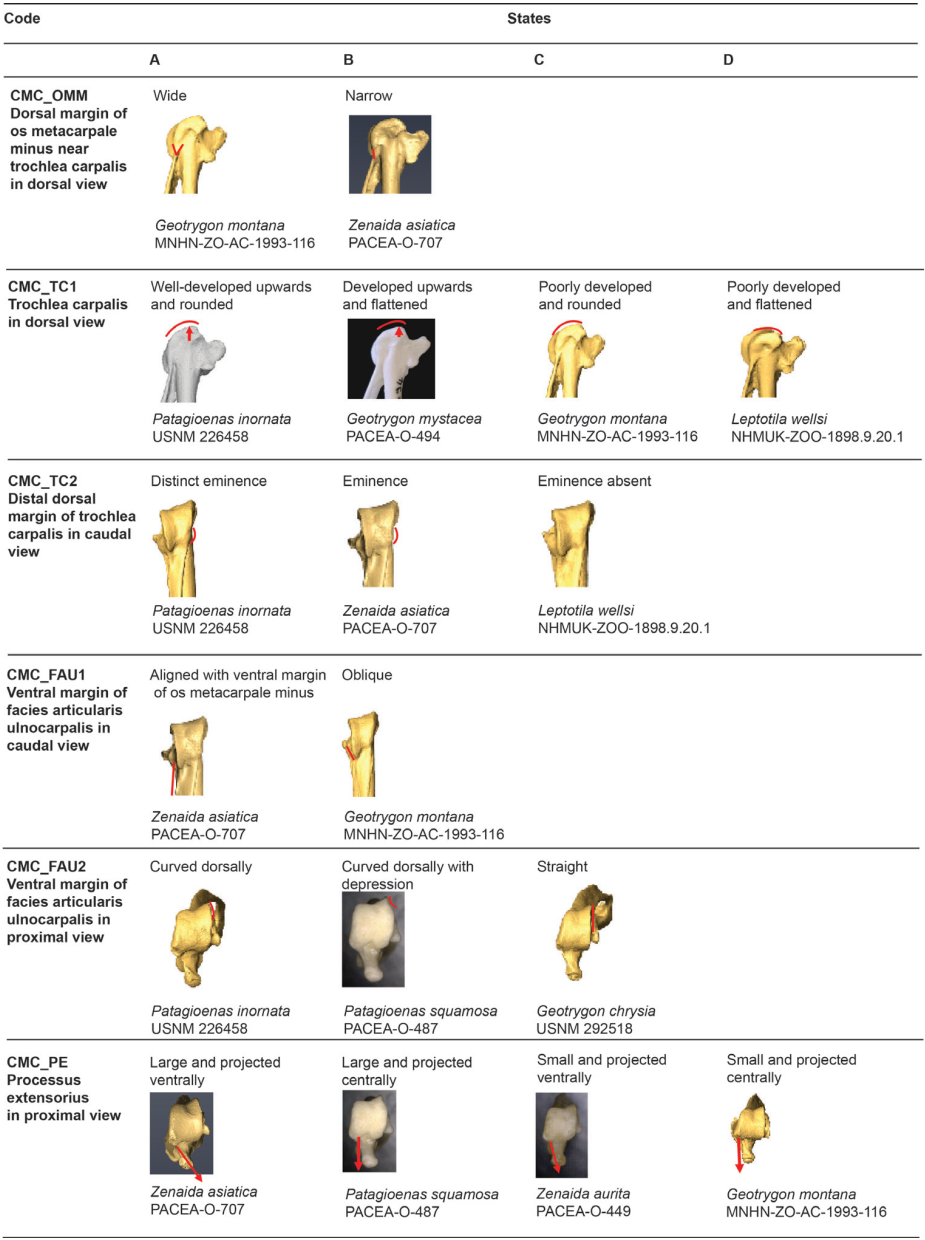
Figure 15, State 1. States of the osteological characters described for the carpometacarpus.
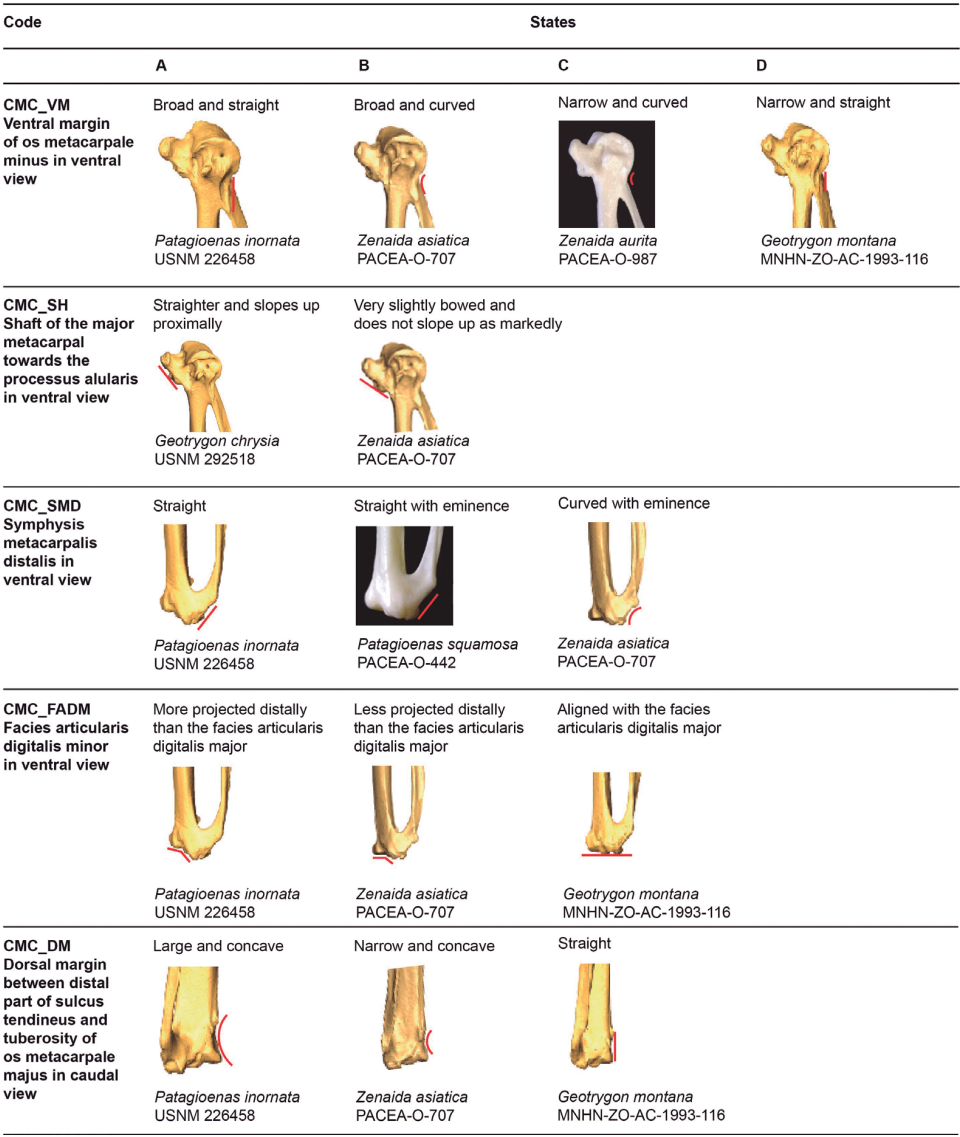
Figure 15, State 2. States of the osteological characters described for the carpometacarpus.
Ventral margin of the facies articularis ulnocarpalis (CMC_FAU1)
Referred material. Zenaida aurita PACEA-O-865 for state A and Geotrygon montana PACEA-O-492 for state B (Table V).
In caudal view, the ventral margin of the facies articularis ulnocarpalis and the ventral margin of the proximal part of the os metacarpale minus are aligned in Zenaida (state A) and P. inornata (Fig. 15, State 1). The ventral margin of the facies articularis ulnocarpalis is oblique compared to the ventral margin of the os metacarpale minus (state B) in Geotrygon, L. wellsi and P. squamosa.
Ventral margin of the facies articularis ulnocarpalis (CMC_FAU2)
Referred material. Patagioenas leucocephala PACEA-O-493 for state A, P. squamosa PACEA-O-487 for state B, Geotrygon montana PACEA-O-492 for state C (Table V).
In proximal view, the ventral margin of the facies articularis ulnocarpalis curves dorsally (state A) in most Patagioenas specimens (Fig. 15, State 1), L. wellsi, Z. auriculata and Z. macroura. In rare cases, a depression is present in state B, as observed in the two P. squamosa specimens.
A straight ventral margin of the facies articularis ulnocarpalis (state C) is characteristic of G. montana and the carpometacarpi of G. mystacea.
Problematic species: no characteristic states are evident in G. chrysia, Z. asiatica and Z. aurita.
Processus extensorius (CMC_PE)
Referred material. Patagioenas squamosa PACEA-O-442 for state A, P. leucocephala PACEA-O-970 for state B, Zenaida aurita PACEA-O-449 for state C, and Z. asiatica PACEA-O-1013 for state D (Table V).
In proximal view, the processus extensorius is large and its caudal extremity projects ventrally (state A) in most of the P. squamosa specimens (Fig. 15, State 1). It is large and oriented centrally (state B) in P. leucocephala and Z. macroura.
A small processus extensorius oriented ventrally (state C) or centrally (state D) is typical of Geotrygon, except for one of the two G. chrysia individuals.
Problematic species: Z. aurita and, to a lesser extent, Z. asiatica and Z. auriculata, present intermediate states or uncharacteristic features.
Ventral margin of the os metacarpale minus (CMC_VM)
Referred material. Patagioenas squamosa PACEA-O-443 for state A, Zenaida asiatica PACEA-O-707 for state B, Geotrygon montana PACEA-O-492 for state C, and Z. aurita PACEA-O-987 for state D (Table V).
In ventral view, the proximal end of the os metacarpale minus has a shallow groove that connects it to the facies articularis ulnocarpalis; the ventral margin of this portion is broad and straight (state A) in P. leucocephala and P. inornata (Fig. 15, State 2).
The ventral margin is curved in Zenaida, broad (state B) in Z. asiatica and Z. macroura, and narrow (state C) in Z. aurita, Z. auriculata. L. wellsi also has a narrow and curve ventral margin.
The carpometacarpi of G. montana and G. mystacea have a narrow, straight ventral margin (state D).
Problematic species: broad ventral margin in P. squamosa and straight ventral margin in G. chrysia.
Shaft of the major metacarpal towards the processus alularis (CMC_SH)
Referred material. Geotrygon chrysia USNM 292518 for state A and Zenaida aurita PACEA-O-865 for state B (Table V).
In ventral view, the character described by Olson and Hilgartner (1982) as distinguishing the two species G. chrysia and Z. aurita can also be used to identify other species. The shaft is straighter and slopes upward proximally in G. chrysia (state A, Fig. 15, State 2), G. mystacea, and P. inornata. The shaft is very slightly bowed and does not slope up as markedly in Z. aurita (state B) and in all other studied species, including G. montana.
Symphysis metacarpalis distalis (CMC_SMD)
Referred material. Geotrygon montana PACEA-O-492 for state A, Patagioenas squamosa PACEA-O-442 for state B and Zenaida asiatica PACEA-O-707 for state C (Table V).
In ventral view, the symphysis metacarpalis distalis is straight (states A-B) in almost all specimens (Fig. 15, State 2; Table V). One feature distinguishing the two states is the presence (state B) or absence of an eminence (state A) on the caudal margin of the os metacarpale minus.
Only a few specimens of P. squamosa, Z. asiatica and Z. aurita have a curved symphysis metacarpalis distalis with an eminence (state C).
Problematic species: no characteristic states in P. leucocephala.
Facies articularis digitalis minor (CMC_FADM)
Referred material. Patagioenas inornata USNM 226458 for state A, P. squamosa PACEA-O-442 for state B, and Geotrygon mystacea PACEA-O-858 for state C (Table V).
In ventral view, the facies articularis digiti minor is more projected distally than the facies articularis digitalis major (state A) in P. inornata (Fig. 15, State 2). These two surfaces are closer (state B) in P. squamosa, P. leucocephala, L. wellsi and almost all specimens of Zenaida. The two surfaces are aligned (state C) in a few Geotrygon and Zenaida specimens.
Problematic species: this character is present in the same proportions as the two states in G. montana and G. chrysia.
Dorsal margin between the distal part of the sulcus tendineus and tuberosity of the os metacarpale majus (CMC_DM)
Referred material. Patagioenas squamosa PACEA-O-442 for state A, Zenaida asiatica PACEA-O-707 for state B, and G. montana PACEA-O-492 for state C (Table V).
In caudal view, the dorsal margin between the distal part of the sulcus tendineus and the tuberosity of the os metacarpale majus is large and concave (state A) in Patagioenas genus (Fig. 15, State 2) and Z. aurita (Table V).
It is narrow and concave (state B) in G. chrysia, L. wellsi, Z. asiatica, Z. auriculata and Z. macroura. It is straight (state C) in most of the G. montana specimens.
Problematic species: G. mystacea present two states.
Carpometacarpus measurement
Carpometacarpus measurements were similar to those provided by Goldberg (1999). In our study, G. mystacea appears smaller rather than similar in size to P. leucocephala, as reported by Steadman et al. (1984b), (Fig. 16). The 5 size ranges of pectoral and wing bones described above are equally evident in the carpometacarpus sample. Several P. leucocephala individuals are similar in size to the other two large pigeons, and Lepotilia falls within the range of small doves. The range of carpometacarpus measurements differs from that of the humerus but is in good agreement with those for the ulna. The length distribution of this element overlaps significantly with G. mystacea and Z. asiatica, G. chrysia, G. montana and, particularly, Z. aurita. However, the length of this bone compared to other elements of the wing appears more discriminating within the group of large doves, in that it distinguishes large specimens of G. mystacea and Z. aurita from G. montana.
Table V. Carpometacarpus characters. Sample size (n) and percentage of states (A-D) and intermediate states (A/B, A/C, B/C, C/D, B/D) for each taxon
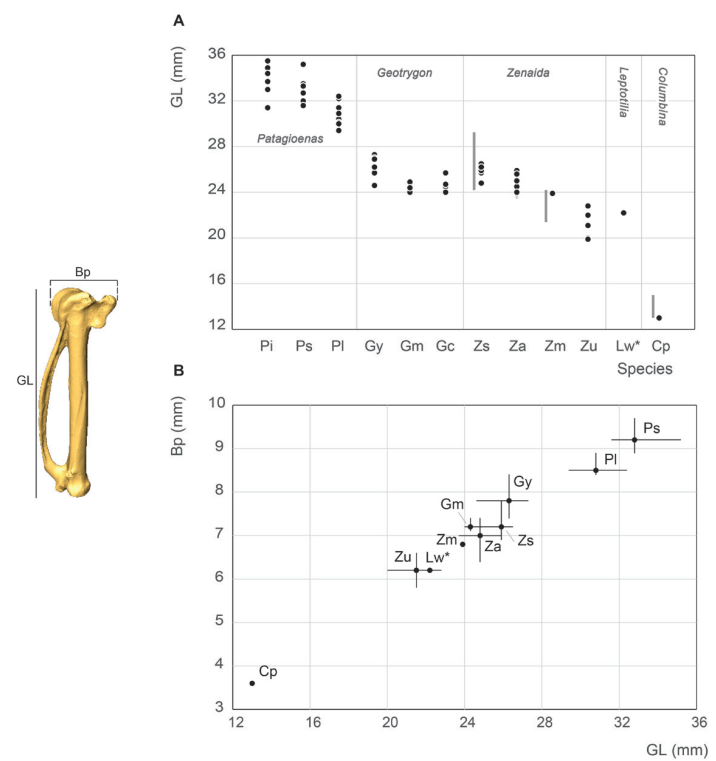
Figure 16. Carpometacarpus measurement in mm: GL, Greatest length (A), Bp, breadth of the proximal end and GL, Greatest length (B). See Appendix C. Abbreviations: Cp, Columbina passerina; Gc, Geotrygon chrysia; Gm, Geotrygon montana; Gy, Geotrygon mystacea; Lw, Leptotila wellsi; Pi, Patagioenas inornata; Pl, Patagioenas leucocephala; Ps, Patagioenas squamosa; Za, Zenaida aurita; Zm, Zenaida macroura; Zs, Zenaida asiatica; Zu, Zenaida auriculata. (*) 3D measurement; the grey vertical line in panel A indicates Goldberg’s measurements (1999).
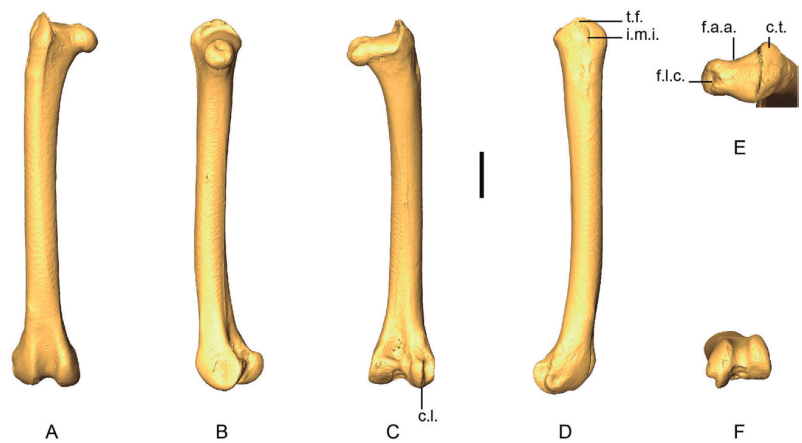
Figure 17. 3D - model of a right femur of Patagioenas inornata USNM 226458 in cranial (A), medial (B), caudal (C), lateral (D), proximal (E), and distal (F), views. Abbreviations: c.l., condylus lateralis; c.t., crista trochanteris; f.a.a., fascies articularis antitrochanterica; f.l.c., fovea ligamentum capitis; i.m.i., impressio musculi iliotrochantericus; t.f., trochanter femoris. Scale bar equals 5 mm.
Femur
Femora from 73 specimens were examined (Appendix P). The overall morphology of this bone reflects the terrestrial mode of life of Geotrygon. Zeinada aurita among Zeinada species has a relatively longer femur. Three osteological criteria of the femur and the features involved in character descriptions are illustrated in figure 17. Two criteria describe the proximal part of the bone and one the distal part (Fig. 18; Table VI).
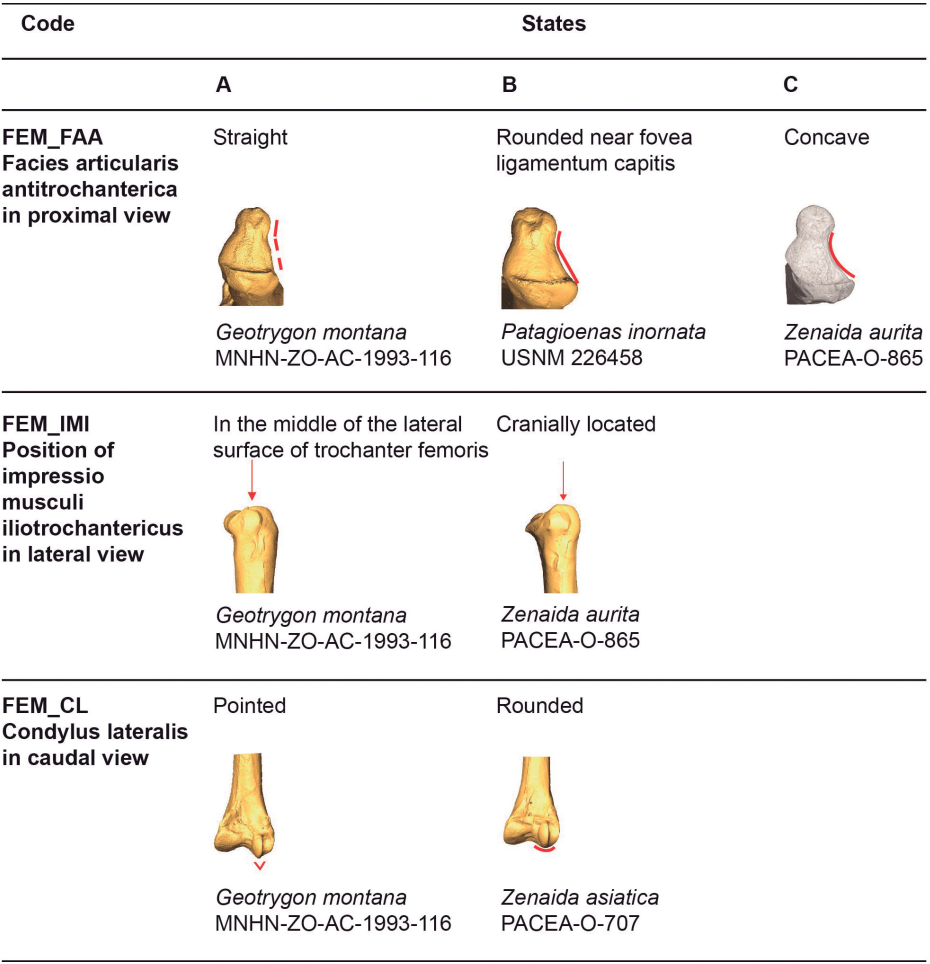
Figure 18. States of the osteological characters described for the femur.
Table VI. Femur characters. Sample size (n) and percentage of states (A-D) and intermediate states (A/B, B/C) for each taxon
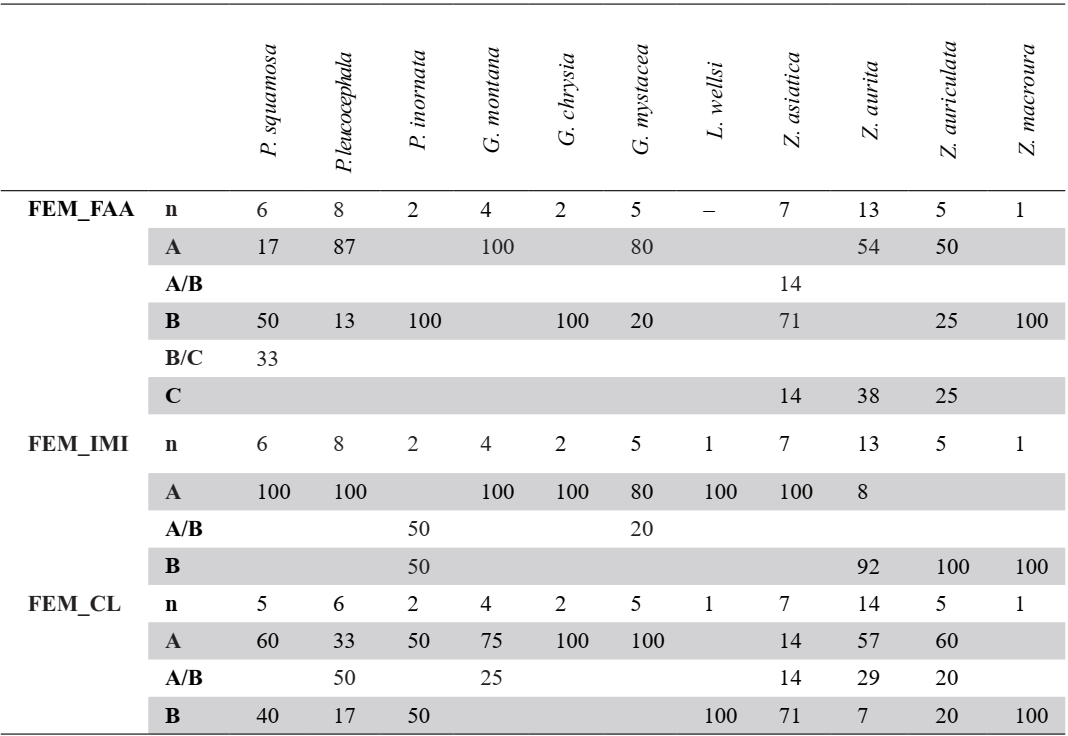
Facies articularis antitrochanterica (FEM_FAA)
Referred material. Patagioenas leucocephala PACEA-O-493 for state A, P. squamosa PACEA-O-442 for state B, and Zenaida aurita PACEA-O-865 for state C (Table VI).
In proximal view, the cranial margin of the facies articularis antitrochanterica is straight (state A) or nearly straight (state B), (Fig. 18). The portion of the facies closest to the fovea ligamentum capitis distinguishes the two states: it is angular in state A as observed in G. montana and G. mystacea and more rounded in state B in G. chrysia. The concave facies articularis antitrochanterica between the crista trochanteris and fovea ligamentum capitis (state C) is a character found only in a very small sample of Zenaida femurs.
Problematic species: several states are observed in P. squamosa, Z. aurita and Z. auriculata.
This character is also not observable in the pathologically-altered femur of L. wellsi.
Position of the impressio musculi iliotrochantericus (FEM_IMI)
Referred material. Patagioenas squamosa PACEA-O-442 for state A and Zenaida aurita PACEA-O-865 for state B.
The impressio musculi iliotrochantericus is most frequently located at the middle of the lateral surface of the trochanter femoris (state A) in Geotrygon, Leptotila and Patagioenas, except for P. inornata (Fig. 18; Table VI). Apart from Z. asiatica, in Zenaida this impressio is more cranially oriented (state B) compared with state A.
Condylus lateralis (FEM_CL)
Referred material. Geotrygon montana PACEA-O-847 for state A and Zenaida asiatica PACEA-O-707 for state B (Table VI).
In caudal view, two states are observable with the condylus lateralis (Fig. 18): pointed (state A) for Geotrygon or rounded (state B) for Leptotila.
The distribution of character states does not distinguish Patagioenas nor Zenaida species.
Femur measurement
Our femur measurements for Z. asiatica and C. passerina do not differ from those reported by Goldberg (1999), (Fig. 19), although the Z. macroura specimen in our sample is larger. The G. montana femur is much longer than those attributed to Z. aurita, a result inconsistent with Steadman et al. (1984b) data for the same species.
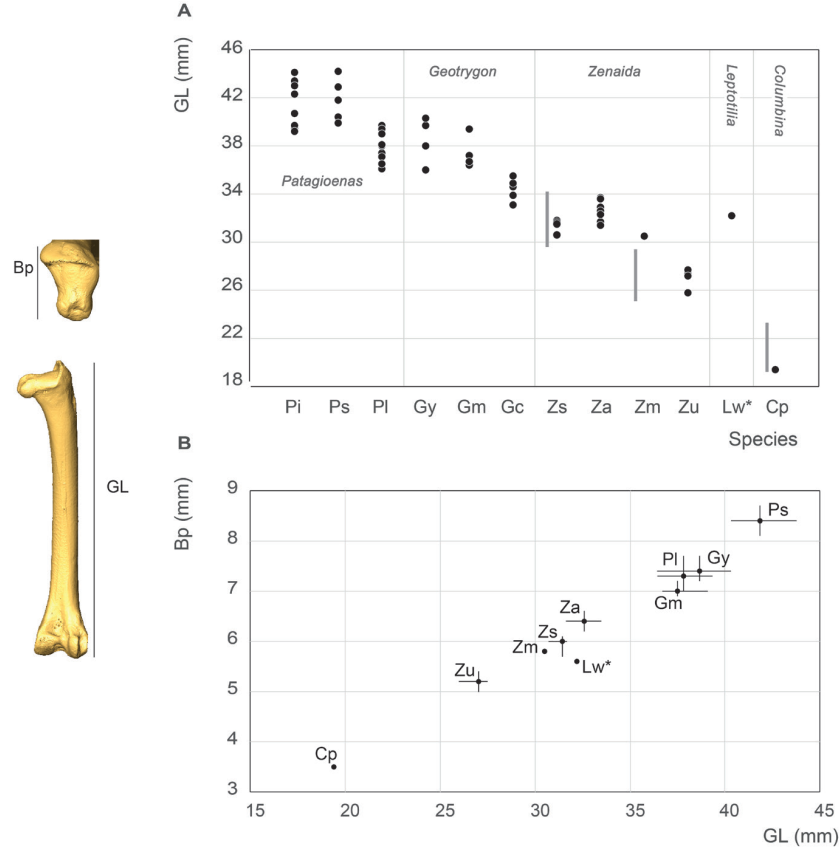
Figure 19. Femur measurement in mm: GL, Greatest length (A), Bp, breadth of the proximal end and GL, Greatest length (B). See Appendix C. Abbreviations: Cp, Columbina passerina; Gc, Geotrygon chrysia; Gm, Geotrygon montana; Gy, Geotrygon mystacea; Lw, Leptotila wellsi; Pi, Patagioenas inornata; Pl, Patagioenas leucocephala; Ps, Patagioenas squamosa; Za, Zenaida aurita; Zm, Zenaida macroura; Zs, Zenaida asiatica; Zu, Zenaida auriculata. (*) 3D measurement; the grey vertical line in panel A indicates Goldberg’s measurements (1999).
P. squamosa and P. inornata fall within the variability of large pigeons. The substantial overlap in femur size between P. leucocephala, G. mystacea, and G. montana makes distinguishing these three taxa based solely on metrtical data difficult, especially given their anatomical similarity. Leptotilia overlaps with the large Zenaida species (Z. asiatica, Appendix Q, and Z. aurita) and G. chrysia with large doves, as observed for the humerus. With that said, the proximal end of Leptotila femur is slenderer than in Zenaida. Our femur measurements group Z. macroua and Z. auriculata in the category of small doves.
Tibiotarsus
Sixty-four tibiotarsi were examined (Appendix R). Complete Patagioenas and Geotrygon tibiotarsi, although difficult to separate based only on maximal length, can be distinguished based on shaft robusticity (Fig. 20; Appendix S). The terrestrial behavior of Geotrygon is reflected in a relatively long tibiotarsus (Baptista et al., 1997) that potentially overlaps with some Patageonias species.
Morphological differences between genera and species are explored based on four osteological criteria, one found in the proximal portion and three in the distal portion of the tibiotarsus (Fig. 21; Table VII).
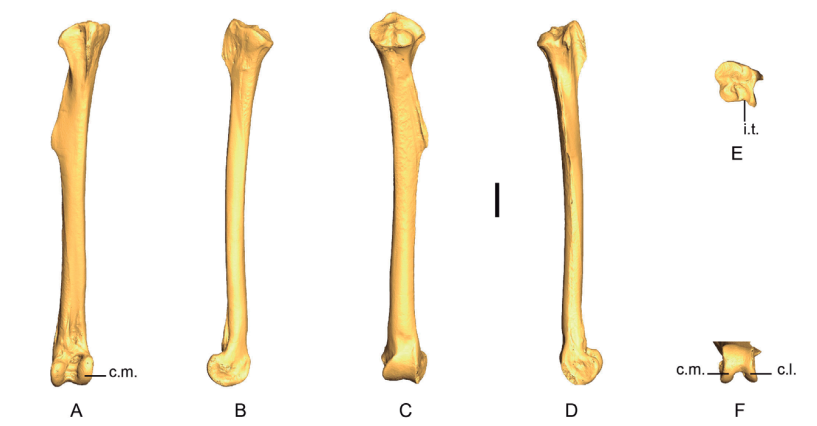
Figure 20. 3D - model of a right tibiotarsus of Patagioenas inornata USNM 290977 in cranial (A), medial (B), caudal (C), lateral (D), proximal (E) and distal (F) views. Abbreviations: c.l., condylus lateralis; c.m., condylus medialis; i.t., incisura tibialis. Scale bar equals 5 mm.
Incisura tibialis (TIB_IT)
Referred material. Zenaida aurita PACEA-O-865 for state A, Patagioenas squamosa PACEA-O-442 for state B (Table VII).
In proximal view, the lateral margin of the incisura tibialis is curved (state A), (Fig. 21) in all species except for P. squamosal, which has a straight incisura tibialis in most specimens (state B).
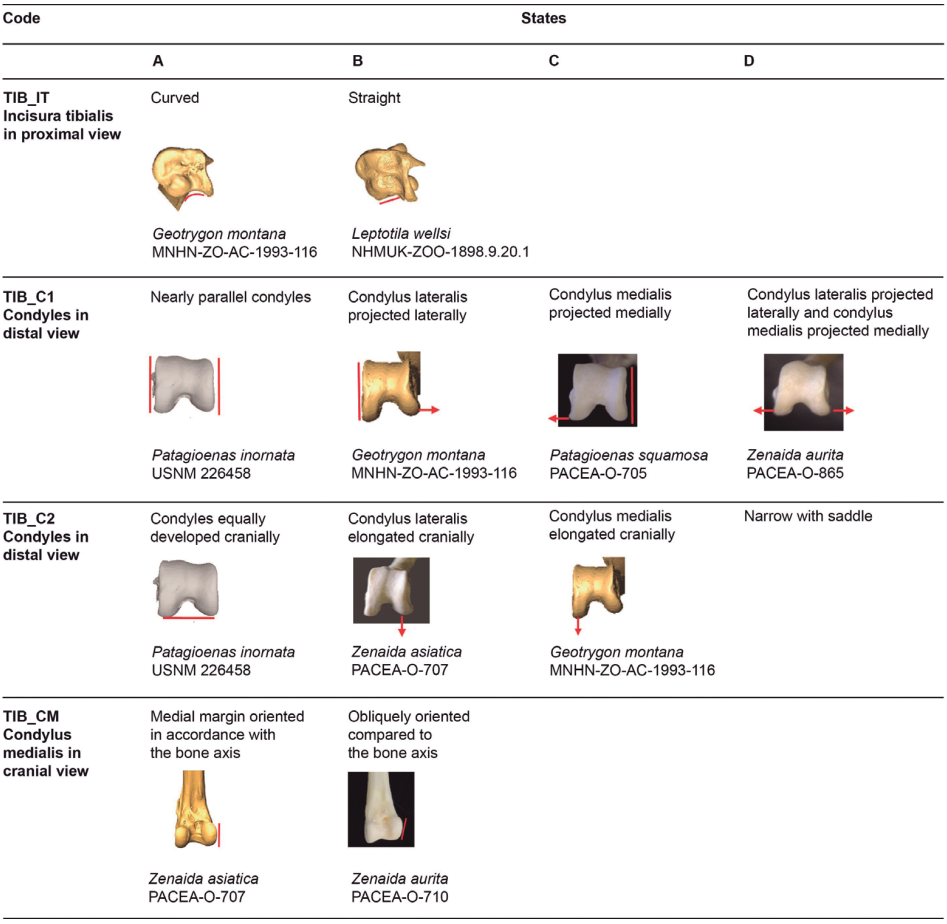
Figure 21. States of the osteological characters described for the tibiotarsus.
Condyles (TIB_C1)
Referred material. Patagioenas inornata USNM 226458 for state A, Geotrygon mystacea PACEA-O-1085 for state B, P. squamosa PACEA-O-705 for state C, and Zenaida aurita PACEA-O-865 for state D (Table VII).
The relative orientation of the condylus lateralis and condylus medialis in distal view distinguish four states; the condyles are nearly parallel (state A), a character observed uniquely in a few Patagioenas specimens, or are non-parallel (states B, C, D), (Fig. 21). In these latter cases, the cranial extremity of the condylus lateralis projects laterally (state B), the cranial extremity of the condylus medialis is oriented medially (state C) or, in most cases (primarily in Z. asiatica), both condyles project in the opposite direction (state D).
Condyles (TIB_C2)
Referred material. Zenaida aurita PACEA-O-865 for state A, Z. asiatica PACEA-O-1004 for state B, and Geotrygon mystacea PACEA-O-494 for state C (Table VII).
In distal view, the cranial extremities of the condyles are equally developed cranially (state A, Fig. 21), especially in P. leucocephala. In specimens when the condyles are not equally developed, the condylus lateralis extends more cranially than the condylus medialis (state B), as is the case with the tibiotarsus of Z. asiatica and Z. auriculata, or conversely (state C), as frequently observed in Geotrygon and the single specimen of L. wellsi.
Table VII. Tibiotarsus characters. Sample size (n) and percentage of states (A-D) and intermediate states (A/B, B/D) for each taxon
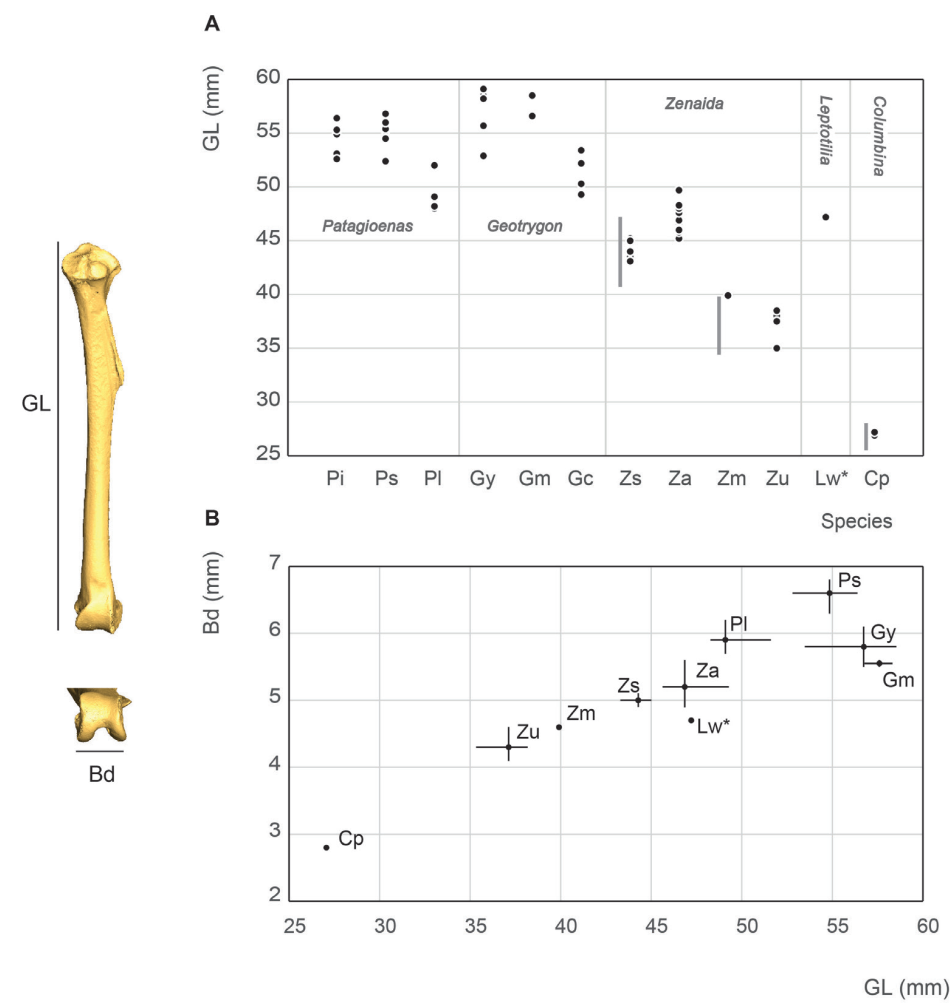
Figure 22. Tibiotarsus measurement in mm: GL, Greatest length (A), Bd, breadth of the distal end and GL, Greatest length (B). See Appendix C. Abbreviations: Cp, Columbina passerina; Gc, Geotrygon chrysia; Gm, Geotrygon montana; Gy, Geotrygon mystacea; Lw, Leptotila wellsi; Pi, Patagioenas inornata; Pl, Patagioenas leucocephala; Ps, Patagioenas squamosa; Za, Zenaida aurita; Zm, Zenaida macroura; Zs, Zenaida asiatica; Zu, Zenaida auriculata. (*) 3D measurement; the grey vertical line in panel A indicates Goldberg’s measurements (1999).
Condylus medialis (TIB_CM)
Referred material. Zenaida asiatica PACEA-O-707 for state A and Z. aurita PACEA-O-865 for state B (Table VII).
In cranial view, the medial margin of the condylus medialis is oriented along the axis of the bone (state A) in almost all Z. asiatica specimens. This character distinguishes this species from the other 3 congeners, which have a tibiotarsus with an oblique condyle (state B), (Fig. 21). This character distinguishes G. mystacea and G. chrysia.
Problematic species: no characteristic states in G. montana.
Tibiotarsus measurement
The Zenaida macroura tibiotarsus measurements are comparable with the largest values reported by Goldberg (1999) while Z. asiatica and C. passerina fall within the variability of North American columbid species (Fig. 22). G. mystacea has a longer tibiotarsus than P. leucocephala (Steadman et al., 1984b), which in turn, is longer than G. montana. Our measurements depict a slighty different pattern; G. montana tibiotarsus length falls within the variability of G. mystacea and both species have a longer tibiotarsus than P. leucocephala. It is also worth noting that the order of the size classes of taxa based on pectoral and wing bones is not the same, primarily due to the greater relative length of Geotrygon tibiotarsus. This leads to the two Lesser Antilles quail-doves (G. mystacea and G. montana) appearing comparable in size to large pigeons, and G. chrysia being the same size as the small pigeon P. leucocephala. However, this pattern reflects Geotrygon’s distinctive slender form, with the GL/Bp ratio setting it apart from other West Indian columbids.
Tarsometatarsus
Tarsometatarsi from 67 specimens have been considered (Appendix T). The comparably longer leg bones of Geotrygon linked to their more terrestrial behavior is even more pronounced with the tarsometatarsus, which had led some to consider the relative gracility of this bone (long and slender) as particular to Geotrygon in comparison to Patagioenas and Zenaida (Steadman et al., 1984b; Olson & Hilgartner, 1982), (Appendix U). Three osteological criteria (2 proximal and 1 distal) and the anatomical features used in the description are illustrated in Figure 23. Among these, the distal character distinguishes Zenaida species (Fig. 24; Table VIII).
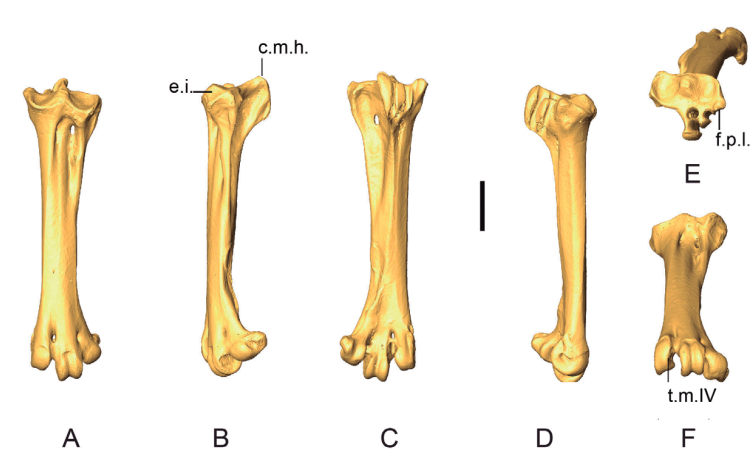
Figure 23. 3D - model of a right tarsometatarsus of Patagioenas inornata USNM 226458 in dorsal (A), medial (B), plantar (C), lateral (D), proximal (E) and distal (F) views. Abbreviations: c.m.h., crista medialis hypotarsi; e.i., eminentia intercotylaris. f.p.l., fossa parahypotarsalis lateralis; t.m.IV., trochlea metatarsi IV. Scale bar equals 5 mm.
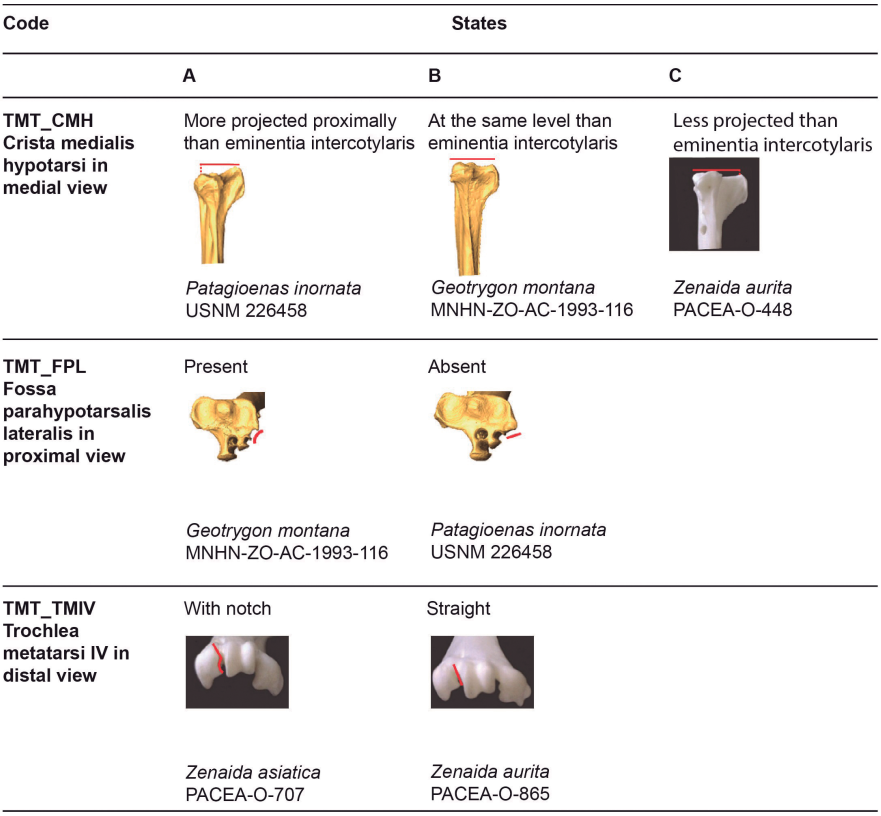
Figure 24. States of the osteological characters described for the tarsometatarsus.
Table VIII. Tarsometatarsus characters. Sample size (n) and percentage of states (A-D) and intermediate states (A/B, B/C) for each taxon
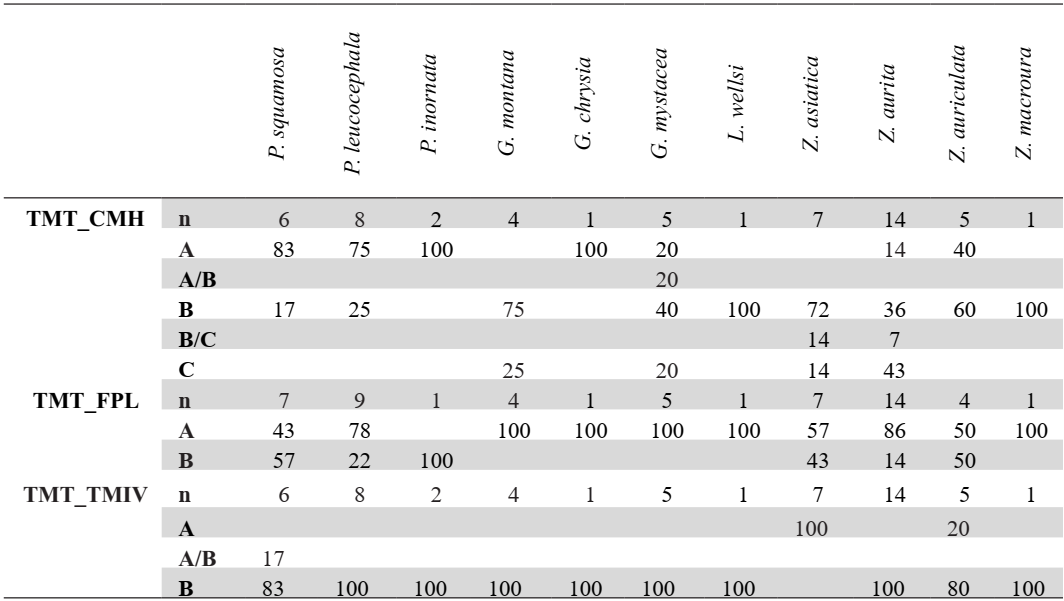
Crista medialis hypotarsi (TMT_CMH)
Referred material. Patagioenas squamosa PACEA-O-442 for state A, Geotrygon montana PACEA-O-492 for state B, and Zenaida aurita PACEA-O-448 for state C (Table VIII).
In medial view, the summit of the crista medialis hypotarsi projects more proximally than the eminentia intercotylaris (state A) in Patagioenas (Fig. 24). Among representatives of Geotrygon, the single G. chrysia specimen exhibits state A while in G. montana the summit of the crista medialis hypotarsi and the eminentia intercotylaris are at the same level (state B). This latter state is also observed in L. wellsi, Z. asiatica and Z. macroura. In some specimens, the crista medialis hypotarsi projects less proximally than the eminentia intercotylaris (state C).
All three states as well as intermediate states are found in G. mystacea and Z. aurita.
Fossa parahypotarsalis lateralis (TMT_FPL)
Referred material. Geotrygon montana PACEA-O-492 for state A and Patagioenas squamosa PACEA-O-443 for state B (Table VIII).
In proximal view, the depth of the fossa parahypotarsalis lateralis varies (state A) in all Geotrygon specimens, most P. leucocephala and Z. aurita, and the single specimens of L. wellsi and Z. macroura (Fig. 24).
The plantar margin of the cotyla lateralis is straight or oblique and the fossa parahypotarsalis lateralis is absent in P. inornata (state B, Fig. 24).
Problematic species: no characteristic states in P. squamosa, Z. asiatica and Z. auriculata.
Trochlea metatarsi IV (TMT_TMIV)
Referred material. Zenaida asiatica PACEA-O-707 for state A and Zenaida aurita PACEA-O-865 for state B (Table VIII).
In distal view, the morphology of the trochlea metatarsi of the fourth digit is characteristic of two states in Zenaida (Fig. 24; Table VIII). In the Z. asiatica, there is a clear notch on the plantar margin of trochlea (state A), while in the other species this margin is straight (state B).
Tarsometatarsus measurement
In our study, length measurements for Zenaida asiatica are comparable with the lowest values reported by Goldberg (1999), (Fig. 25). Our Z. macroura specimen is substantially longer than those in Goldberg’s (1999) study. G. montana is much longer than G. mystacea compared to the observations of Steadman et al. (1984b). The comparably longer leg bones of the tarsometatarsus in columbid genera with more terrestrial behavior is more important than seen in the other leg bones, with Geotrygon specimens having the longest values. Lower length values for G. chrysia overlap with the higher value of the large pigeons P. inornata and P. squamosa. The two large Zenaida and Leptotilia fall within the range of Patagioenas. Similir to the tibiotarsus, this overlap reflects the more slender form of Geotrygon and Leptotilia. Unlike the tibiotarsus, this more gracile aspect, although slight, is neverless more evident in Zenaida than Patagioeonas.
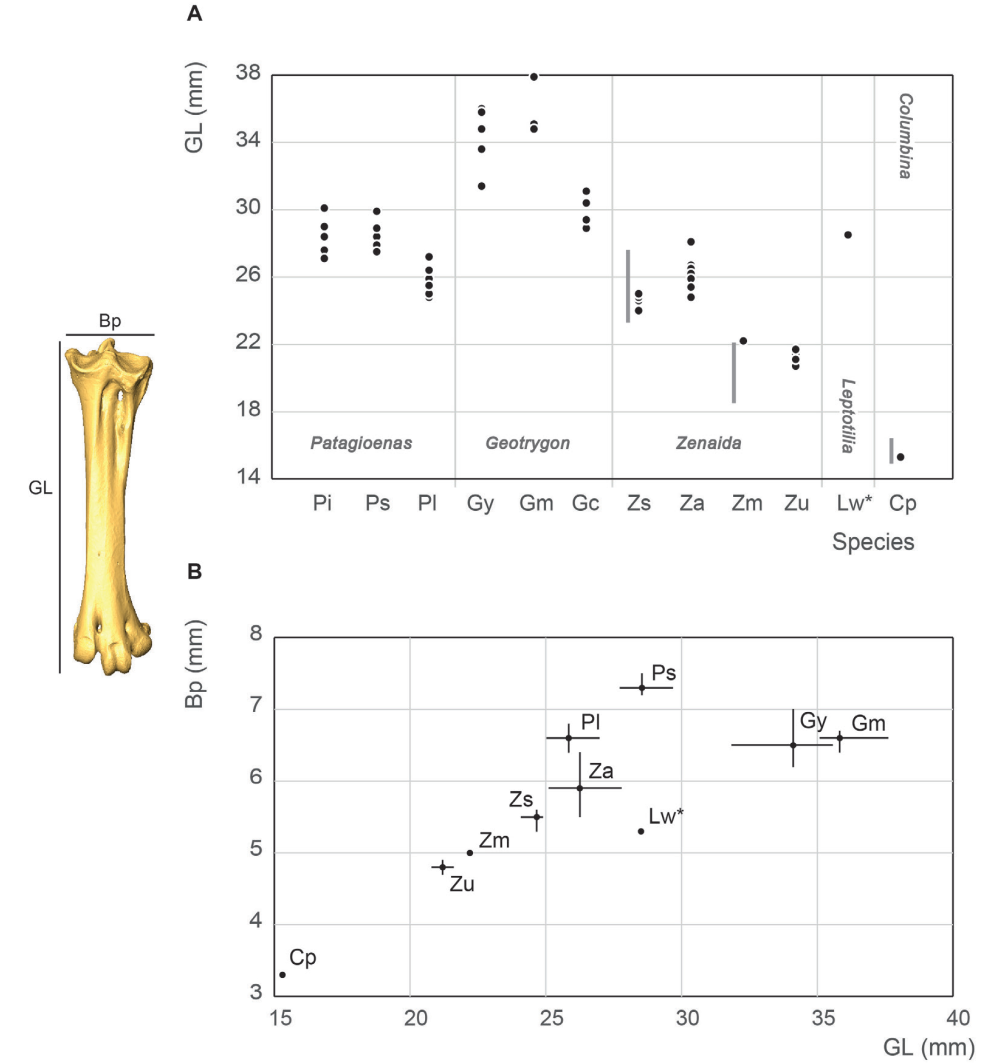
Figure 25. Tarsometatarsus measurement in mm: GL, Greatest length (A), Bp, breadth of the proximal end and GL, Greatest length (B). See Appendix C. Abbreviations: Cp, Columbina passerina; Gc, Geotrygon chrysia; Gm, Geotrygon montana; Gy, Geotrygon mystacea; Lw, Leptotila wellsi; Pi, Patagioenas inornata; Pl, Patagioenas leucocephala; Ps, Patagioenas squamosa; Za, Zenaida aurita; Zm, Zenaida macroura; Zs, Zenaida asiatica; Zu, Zenaida auriculata. (*) 3D measurement; the grey vertical line in panel A indicates Goldberg’s measurements (1999).

DISCUSSION
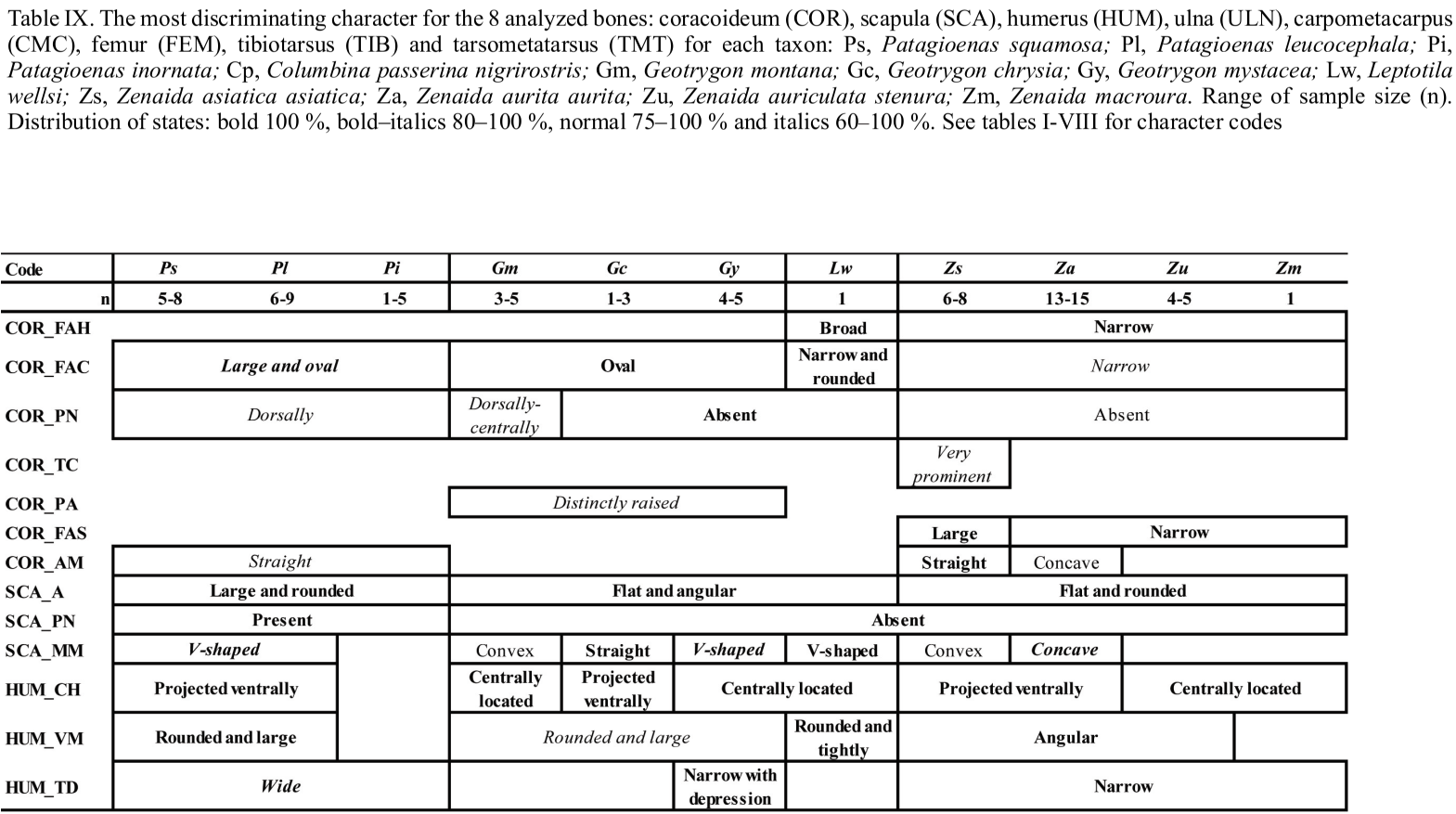
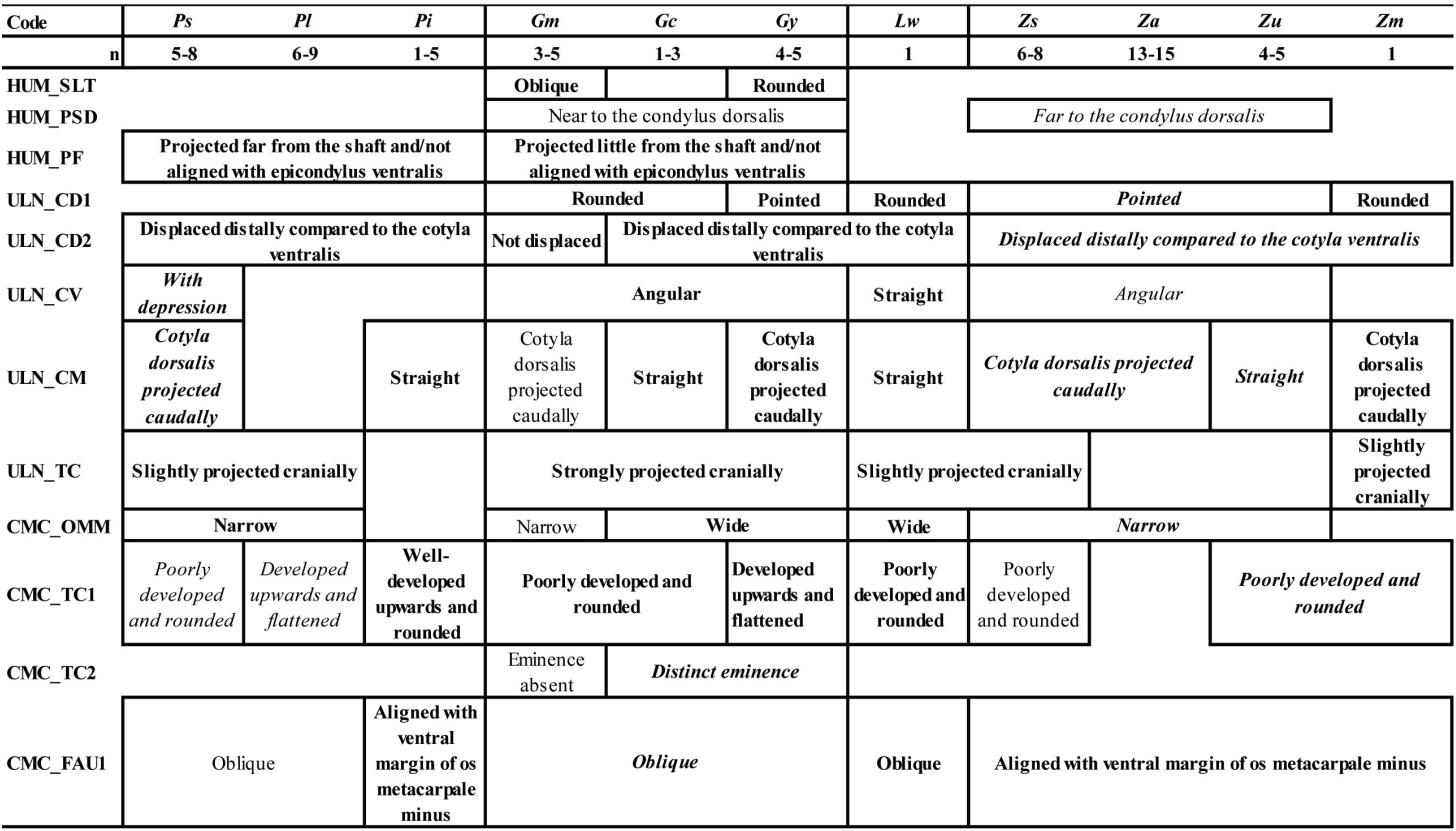
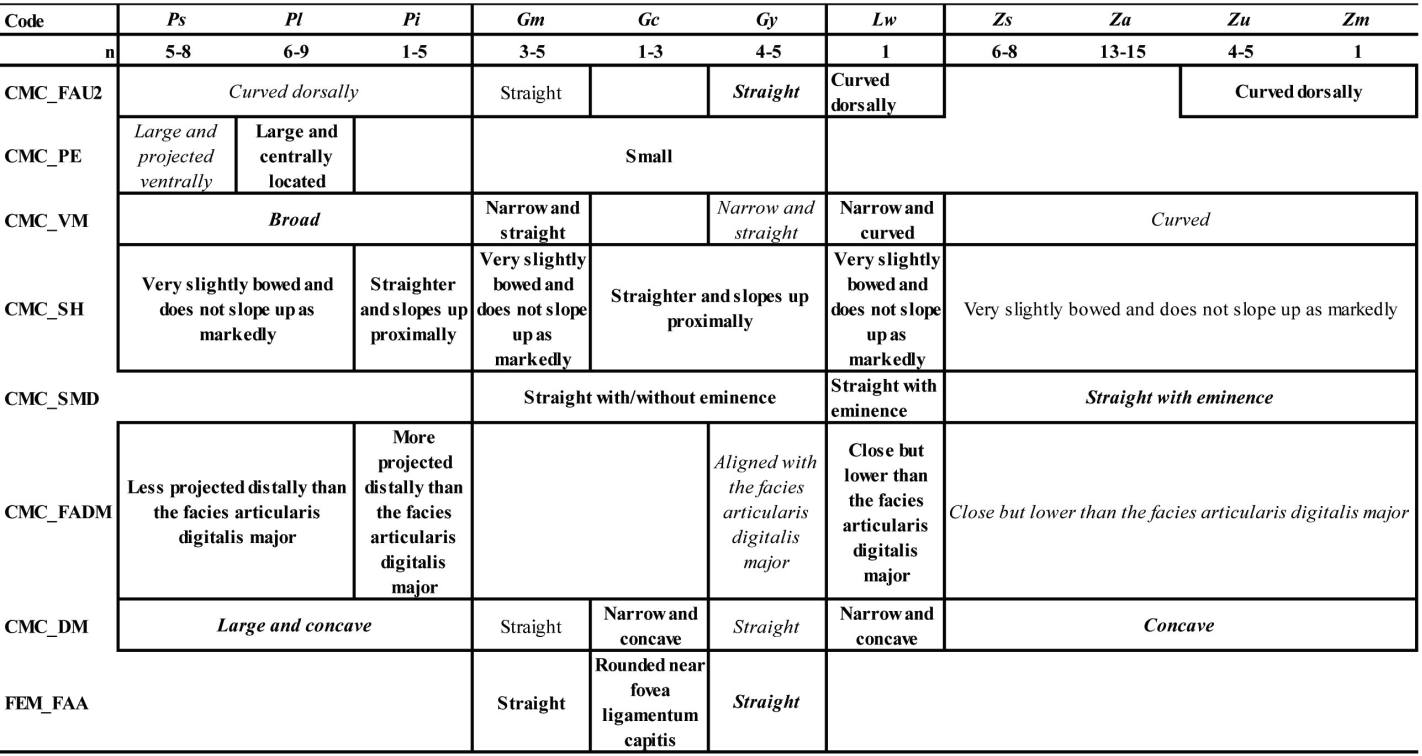
A comparison of osteological and biometrical data for the 8 bones of the upper and lower limbs and pectoral girdle of pigeons and doves demonstrate the 12 taxa in our analysis to differ in several morphological traits and/or in size. The most frequently recorded states of the 42 osteological characters described for the eight analyzed postcranial skeleton elements are available as Table IX.
Two characters evident on elements of the pectoral girdle (COR_FAC and SCA_A) distinguish specimens of the four genera in our sample (Patagioenas, Geotrygon, Zenaida and Leptotila). Several wing bone characters equally separate Patagioenas and Geotrygon (HUM_VM) and Geotrygon and Zenaida (ULN_CV). One character state of the coracoideum (COR_PN), in addition to being present in Patagioenas, is also exhibited in one Geotrygon species. Six characters of the coracoideum, humerus, and carpometacarpus are distinguishing characteristics separating Patagioenas from Geotrygon (HUM_PF), Patagioenas from Zenaida (HUM_TD), Geotrygon from Zenaida (HUM_VM, HUM_PSD and CMC_FAU1), and Zenaida from Leptotila (COR_FAH).
Apart from the humerus, seven bones separate the different genera: Patagioenas (COR_AM, SCA_PN and TMT_CMH), Geotrygon (COR_PA, ULN_TC, FEM_CL, TIB_C2 and TMT_FPL) and Zenaida (CMC_SH and CMC_SMD). Four bones lend themselves for identification to species level: P. squamosa (TIB_IT), P. leucocephala (CMC_PE), G. mystacea (CMC_FADM), G. montana (COR_PN and ULN_CD2) and Z. asiatica (COR_TC, TIB_C1 and TMT_TMIV).
In addition, the absence of some character states can help in identifying columbid bones and distinguish between different genera: Patagioenas (SCA_MM, HUM_PF, CMC_FAU2, CMC_FADM, CMC_DM and TMT_CMH), Geotrygon (COR_TC, HUM_SLT, HUM_PF,
CMC_PE, CMC_SMD, CMC_DM, FEM_FAA) and Zenaida (SCA_MM, CMC_VM and CMC_ FADM). Patagioenas and Geotrygon both share two characters (CMC_TC1 and TIB_C2).
Interspecific differences can be observed in three genera. One character of the carpometacarpus and two of the humerus distinguish three Patagioenas species (HUM_ CH, HUM_VM and CMC_TC1). On the other hand, multiple characters reliably separate Geotrygon species: G. chrysia differs from the congeneric species in five characters (HUM_CH, ULN_CM, CMC_DM, FEM_FAA and TIB_C1); G. montana differs in two characters on the carpometacarpus (CMC_TC2 and CMC_OMM) and G. mystacea in two characters (ULN_CD1 and CMC_TC1). One character on the humerus distinguishes G. mystacea from G. montana (HUM_SLT). Three characters found on the coracoideum, femur and tibiotarsus distinguish Z. asiatica from congeneric species (COR_FAS, FEM_IMI and TIB_CM) or from Z. aurita (COR_AM). Two characters on the humerus (HUM_CH) and carpometacarpus (CMC_FAU2) distinguish Z. asiatica and Z aurita from Z. auriculata and Z. macroura.
Overall, our measurements evince five size categories: large (P. inornata and P. squamosa) and small pigeons (P. leucocephala), large (Geotrygon spp., Z. asiatica and Z. aurita), medium (Z. macroura and Z. auriculata), and small doves (C. passerina). Independent of the skeletal element considered, Columbina passerina is always the smallest species and no overlap in size can be observed with the other Caribean columbids. The position of the single L. wellsi specimen amongst medium or large doves varies between skeletal elements, indicating the need for additional data for this species. More specifically, this size ordering changes depending on body size, which varies as a function of the terrestrial habits of birds (Baptista et al., 1997). Hence, the size and shape of complete pectoral (coracoideum and scapula) and wing bones (humerus, ulna and carpometacarpus) reliably separate large and small pigeons from large and medium doves. For wing bones, P. leucocephala occupies an intermediate position between the large pigeons and the quail-doves, the latter being grouped with the large Zenaida (Z. asiatica and Z. aurita). The relative position of the Geotrygon changes according to the leg bones considered; the two large pigeons generally have longer femurs than G. mystacea and G. montana, with P. leucocephala grouped with the latter. The opposite is true for the tarsometatarsus, with the tibiotarsus occupying an intermediate position. Clear differences are also evident between these limb elements in terms of robusticty. The allometry evident between wing and leg bones of New World quail-doves (Geotrygon) compared to the other genera potentially reflects an adaptation to terrestrial behavior (Wetmore, 1922; Baptista et al., 1997).
When compared with data provided by Goldberg (1999), several minor metrical differences emerge. The place of Geotrygon within Antillean columbids in our study also differs from that reported by Steadman et al. (1984b), except for the femur and tarsometatarsus. This discrepancy potentially reflects different populations in the two samples, as our Geotrygon mystacea and G. montana specimens come uniquely from the Guadeloupe Islands while the Smithsonian Institute G. montana sample used by Steadman et al. (op. cit.) includes numerous continental specimens. These difference could also reflect a limited sample that underestimates intra-specific variability.
The precise origin of each specimen in this study (Appendix A) and associated metric data (Appendices C, D, G, I, L, N, P, R, T) can be integrated in future studies to better identify potential regional variability in Columbidae.
CONCLUSIONS
The morphometric analysis of the postcranial skeleton of the main Caribbean pigeons and doves demonstrates identifying avian remains to species can be difficult even with a large modern osteological sample. The five size classes identified and 42 diagnostic characters described in our study reliably distinguish skeletal elements of different columbid species. Combining several characters and measurements on complete bones will undoubtedly help in the determination of columbid remains recovered from archaeological and paleontological deposits, especially those from sites excavated in the French West Indies over the past 60 years.
ACKNOWLEDGEMENTS
Funding to this study was provided by the ECSIT Project: “Écosystèmes insulaires tropicaux, réponse de la faune indigène terrestre de Guadeloupe à 6 000 ans d’anthropisation du milieu”, conducted by the CNRS with a financial support from the European PO-FEDER program (grant n° 2016-FED-503), the Guadeloupe Regional Council, the DAC of Guadeloupe (PCR Extinctions), and the CNRS (SEEG ECSIT). Many thanks are due to the curators and staff of the different museums: A. Kratter, J-L. Bernos, S. Guimarães, C. Lefèvre, C. Milensky, D. Steadman, J. White and T. Webber. Thanks are also due to M. Bessou, D. Cochard, C. Haget, R. Ledevin, E. Pubert and S. Renou for their technical support; V. Conche and N. Jeantet for the 3D modeling. We would also like to thank Anna Rufà Bonache for the Spanish translation of the abstract and Brad Gravina for the English editing and constructive comments. We also thank the editor and two anonymous reviewers who provided constructive feedback and comments on this paper and J. Orihuela for his bibliographical help concerning the Cuban material.
Literatura Citada
Arredondo, O. 1984. Sinopsis de las aves halladas en depósitos fasilíferos Pleisto-Holocénicos de Cuba. Reporte de Investigación del Instituto de Zoología, 17: 1-35.
Baptista, L. F., P. W. Trail, & H. M. Horblit. 1997. Family Columbidae (pigeons and doves), (pp. 60-243) In: del Hoyo, J., A. Elliott & J. Sargatal (Eds.). Handbook of the birds of the world. Vol. 4: Sandgrouse to cuckoos. Lynx Edicions, Barcelona.
Baumel, J. J., & L. M. Witmer. 1993. Osteologia (pp. 45-132). In: Baumel, J. J., A. S. King, J. E. Breazile, H. E. Evans & J. C. Vanden Berge (Eds.). Handbook of Avian Anatomy: Nomina Anatomica Avium, Publications of the Nuttall Ornithological Club, NO. 23, Cambridge.
Brodkorb, P. 1959. Pleistocene birds from New Providence Island, Bahamas. Bulletin of the Florida State Museum, Biological Sciences, 4: 349-371.
Driesch von den, A. 1976. A guide to the measurements of animal bones from archaeological sites. Peabody Museum Bulletin, Vol. 1. Peabody Museum of Archaeology and Ethnology, Harvard University, Cambridge, 138 pp.
Fick, O. K. W. 1974. Vergleichend morphologische Untersuchungen an Einzelknochen europäischer Taubenarten. Inaugural-Dissertation, Ludwig-Maximilians-Universität, München, 93 pp.
Gibbs, D., E. Barnes, & J. Cox. 2001. Pigeons and doves: a guide to the pigeons and doves of the world, Yale University Press, New Haven, CT, 616 pp.
Goldberg, P. M. 1999. Statistical comparisons of the osteology of Southwestern Columbidae. Journal of archaeological science, 26 (12): 1459-1471.
Grouard, S. 2001. Subsistance, systèmes techniques et gestion territoriale en milieu insulaire antillais précolombien: exploitation des vertébrés et des crustacés aux époques Saladoïdes et Troumassoïdes de Guadeloupe (400 av. JC à 1500 ap. JC). Dissertation presented for the Graduate School, University of Paris.
Grouard, S. 2013. Chasses, pêches et captures de faunes vertébrées et crustacées des occupations côtières récentes du Sud de la Martinique (Saladoïde récent, Vè siècle Ap. J.-C. - Suazoïde récent, XVe s. ap. J.-C.) (pp. 115-161). In: Bérard B. (Ed.). Martinique, terre amérindienne. Une approche pluridisciplinaire. Sidestone Press, Leiden.
Jiménez-Vázquez, O. 2001. Registros ornitológicos en residuarios de dieta de los aborígenes precerámicos cubanos. El Pitirre, 14: 120-126.
Jiménez-Vázquez, O., & R. Arrazcaeta. 2008. Las aves en la arqueología histórica de La Habana Vieja. Boletín del Gabinete de Arqueología, 7: 17-29.
Jiménez-Vázquez, O., & R. Arrazcaeta. 2015. Las aves y su relación con las culturas precolombinas de Cuba. Boletín del Gabinete de Arqueología, 11: 141-157.
Livezey, B. C., & R. L. Zusi. 2006. Phylogeny of Neornithes. Bulletin of Carnegie Museum of Natural History, 37: 1-544.
Newsom, L. A., & E. S. Wing. 2004. On land and sea: Native American uses of biological resources in the West Indies. University of Alabama Press, Tuscaloosa, 344 pp.
Olson, S. L. 2011. The fossil record and history of doves on Bermuda (Aves: Columbidae). Proceedings of the Biological Society of Washington, 124: 1-6.
Olson, S. L., & W. B. Hilgartner. 1982. Fossil and subfossil birds from the Bahamas. In S. L. Olson, (Ed.). Fossil vertebrates from the Bahamas. Smithsonian Institution Press, Washington. Smithsonian Contributions to Paleobiology, 48: 25-55.
Olson, S. L., & J. W. Wiley. 2016. The Blue-headed Quail-Dove (Starnoenas cyanocephala): an Australasian dove marooned in Cuba. The Wilson Journal of Ornithology, 128: 1-21.
Orihuela, J., L. Pérez Orozco, J. L. Álvarez Licourt, R. A. Viera Muñoz, & C. Santana Barani, 2020. Late Holocene land vertebrate fauna from Cueva de los Nesofontes, Western Cuba: stratigraphy, last appearance dates, diversity and paleoecology. Palaeontologia Electronica 23 (3): a57: doi: 10.26879/995.
Pregill, G. K., D. W. Steadman, & D. R. Watters. 1994. Late quaternary vertebrate faunas of the Lesser Antilles: historical components of Caribbean biogeography, 51 pp.
Raffaele, H. A., J. Wiley, O. H. Garrido, A. Keith, & J. I. Raffaele. 2003. Birds of the West Indies, Princeton University Press, Princeton, NJ, 231 pp.
Steadman, D. W., & J. Franklin. 2020. Bird populations and species lost to Late Quaternary environmental change and human impact in the Bahamas. Proceedings of the National Academy of Sciences, 117: 26833-26841.
Steadman, D. W., G. K. Pregill, & S. L. Olson. 1984a. Fossil vertebrates from Antigua, Lesser Antilles: Evidence for late Holocene human-caused extinctions in the West Indies. Proceedings of the National Academy of Sciences, 81: 4448-4451.
Steadman, D. W., D. R. Watters, E. J. Reitz, & G. K. Pregill. 1984b. Vertebrates from archaeological sites on Montserrat, West Indies. Annals of the Carnegie Museum of Natural History, 53: 1-29.
Tellkamp, M. P. 2005. Prehistoric exploitation and biogeography of birds in coastal and Andean Ecuador. Dissertation presented for the Graduate School, University of Florida.
Tomek, T., & Z. M. Bochenski. 2009. A key for the identification of domestic bird bones in Europe: Galliformes and Columbiformes, Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Krakow, 111 pp.
Van der Klift, H. M. 1992. Faunal remains of the Golden Rock site (pp. 74-84). In: Versteeg A. H., & K. Schinkel (Eds.). The archaeology of ST. Eustatius, the Golden Rock site. Publication of the Foundation for Scientific Research in the Caribbean Region, Amsterdam, The Netherlands.
Wells, J., & A. Wells. 2001. Pigeons and Doves (pp. 319-325). In: Elphick, C., J. Dunning, & D. Sibley (Eds.). The Sibley Guide to Bird Life and Behavior. New York: Alfred A. Knopf.
Wetmore, A. 1920. Five new species of birds from cave deposits in Porto Rico. Proceedings of the Biological Society of Washington, 33: 77-82.
Wetmore, A. 1922a. Bird remains from the caves of Porto Rico. Bulletin of the American Museum of Natural History, 46: 297-333.
Wetmore, A. 1922b. Remains of birds from caves in the republic of Haiti. Smithsonian Miscellaneaous Collections, 74: 1-4.
Wetmore, A. 1925. Another record of birds from the Island of St. Croix. The Auk, 42 (3): 446.
Wetmore, A. 1937. Bird remains from cave deposits on Great Exuma Island in the Bahamas. Bulletin of the Museum of Comparative Zoology, 80: 427-441.
Wiley, J. W. 1991. Ecology and behavior of the Zenaida Dove. Ornitologia Neotropical, 2 (2): 49-75.
Worthy, T. H. 2001. A giant flightless pigeon gen. et sp. nov. and a new species of Ducula (Aves: Columbidae), from Quaternary deposits in Fiji. Journal of the Royal Society of New Zealand, 31 (4): 763-794.
APPENDIX A
Modern comparative specimens. (F) female and (M) male. Unavailable data is indicated by a dash, 3D-models with an asterisk
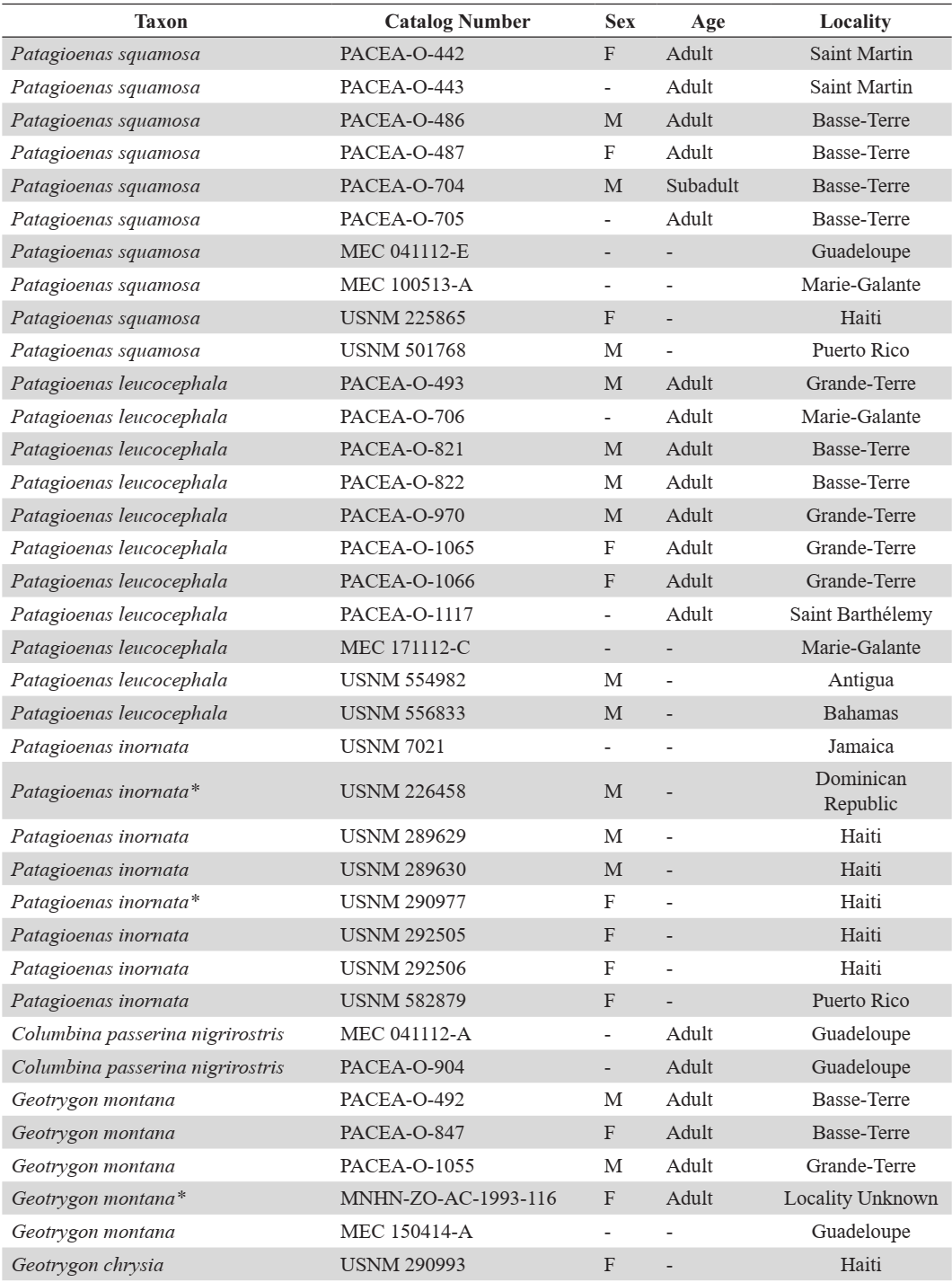
APPENDIX B
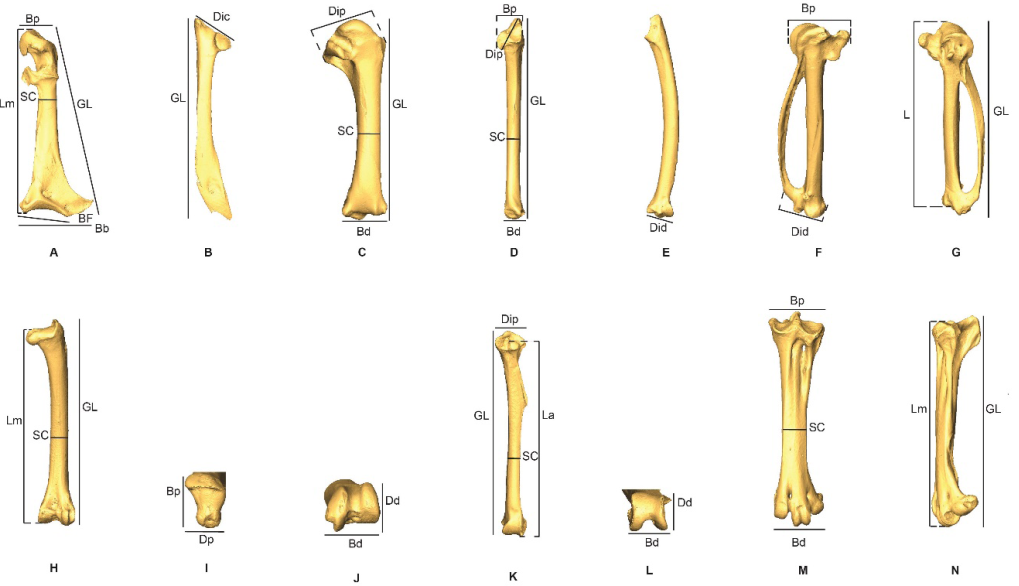
Measurements of the Columbidae modified from von den Driesch (1976). A, coracoideum in dorsal view; B, scapula in lateral view; C, humerus in caudal view; D-E, ulna in cranial and ventral views; F-G, carpometacarpus in dorsal and ventral views; H-J, femur in caudal, proximal and distal views; K-L, tibiotarsus in caudal and distal views; M-N, tarsometatarsus in dorsal and medial views. Abbreviations: Bb, (greatest) basal breadth; Bd, breadth of the distal end; BF, breadth of facies articularis basalis; Bp, breadth of the proximal end; Dd, depth of the distal end; Dic, (greatest) cranial diagonal; Did diagonal of the distal end; Dp, (greatest) depth of the proximal end; Dip, (greatest) diagonal of the proximal end; GL, greatest length; L length of the metacarpus II; La, axial length; Lm, medial length; Sc, smallest breadth of the corpus.
APPENDIX C
Measurements (in mm) for Columbidae. Unavailable data is indicated by a dash, 3D models with an asterisk. See Appendix B for significance of the measurements
APPENDIX D
Measurements (in mm) and characters of coracoideum from modern specimens (character states see Fig. 2). Unavailable data is indicated by a dash, 3D-models with an asterisk
APPENDIX E
3D - models of coracoidea in dorsal view. A, Patagioenas inornata USNM 226458; B, Geotrygon montana MNHN-ZOAC-1993-116; C, Leptotila wellsi NHMUK-ZOO-1898.9.20.1; D, Zenaida asiatica PACEA-O-707. A-B, D, right side; C, left side. Scale bar equals 5 mm.
APPENDIX F
Additional characters of the coracoideum.
The following omal characters allow distinguish Geotrygon from Zenaida.
In agreement with Olson and Hilgartner, in G. chrysia, the glenoid facet (facies articularis humeralis) in lateral view is smaller than in Z. aurita and the “scapular facet is not situated as far sternally on the shaft as Z. aurita” (Olson & Hilgartner,1982 p. 35).
Two characters (shape of cotyla scapularis) are unreliable for identifying specimens to species due to the substantial variability of Geotrygon and Zenaida.
In dorsal view, the shape of cotyla scapularis is triangular in Geotrygon and uncharacteristic in Zenaida (irregular form, rounded or triangular).
In medial view, the cotyla scapularis is most often rounded in G. montana and straight in Z. asiatica and some individuals of Z. aurita.
In addition, the 2 distal characters are situated in the area of the facies articularis sternalis and the angulus medialis. The facies articularis sternalis of G. montana in sternal view is triangular in 60 % of specimens.
In medial view, the angulus medialis of Z. asiatica is often large with a symmetric rhomboid shape.
LITERATURE CITED
Olson, S. L., & W. B. Hilgartner. 1982. Fossil and subfossil birds from the Bahamas (25-55). In: Olson, S. L. (Ed.). Fossil vertebrates from the Bahamas. Smithsonian Institution Press, Washington D.C., Smithsonian Contributions to Paleobiology, 48.
APPENDIX G
Measurements (in mm) and characters of scapula from modern specimens (character states see Fig. 6). Unavailable data is indicated by a dash, 3D-models with an asterisk

APPENDIX H
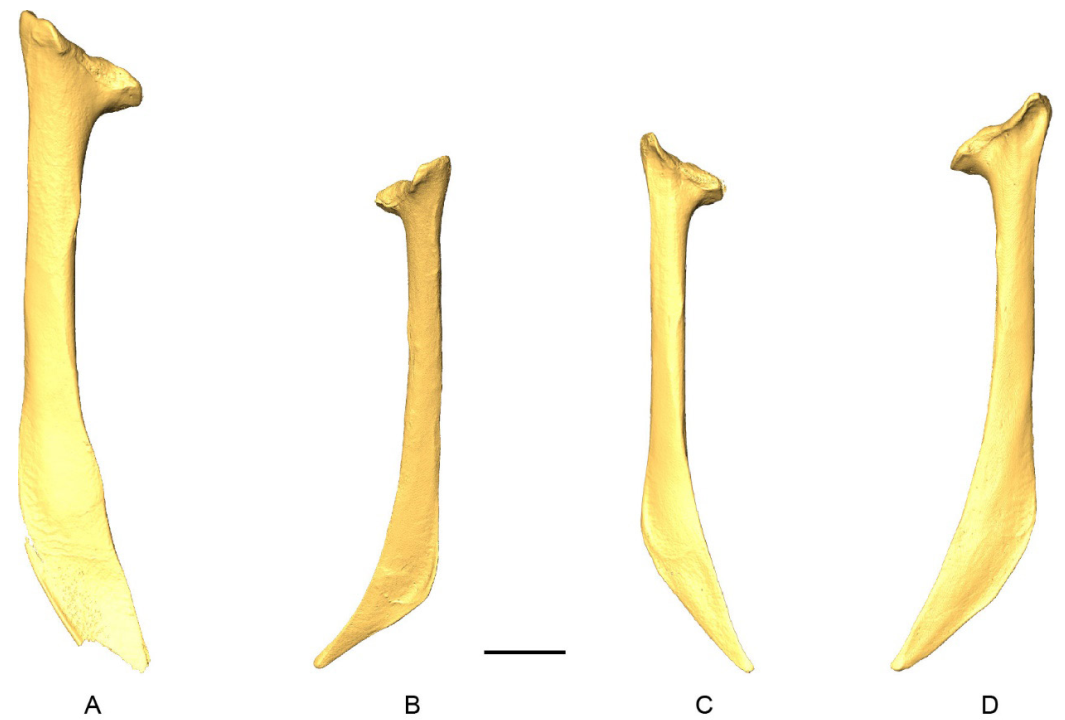
D-models of scapulae in medial view. 3 A, Patagioenas inornata USNM 290977; B, Geotrygon montana MNHN-ZO-AC-1993-116; C, Leptotila wellsi NHMUK-ZOO-1898.9.20.1; D, Zenaida asiatica PACEA-O-707. B, D, right side; A, C, left side. Scale bar equals 5 mm.
APENDIX I
APPENDIX J
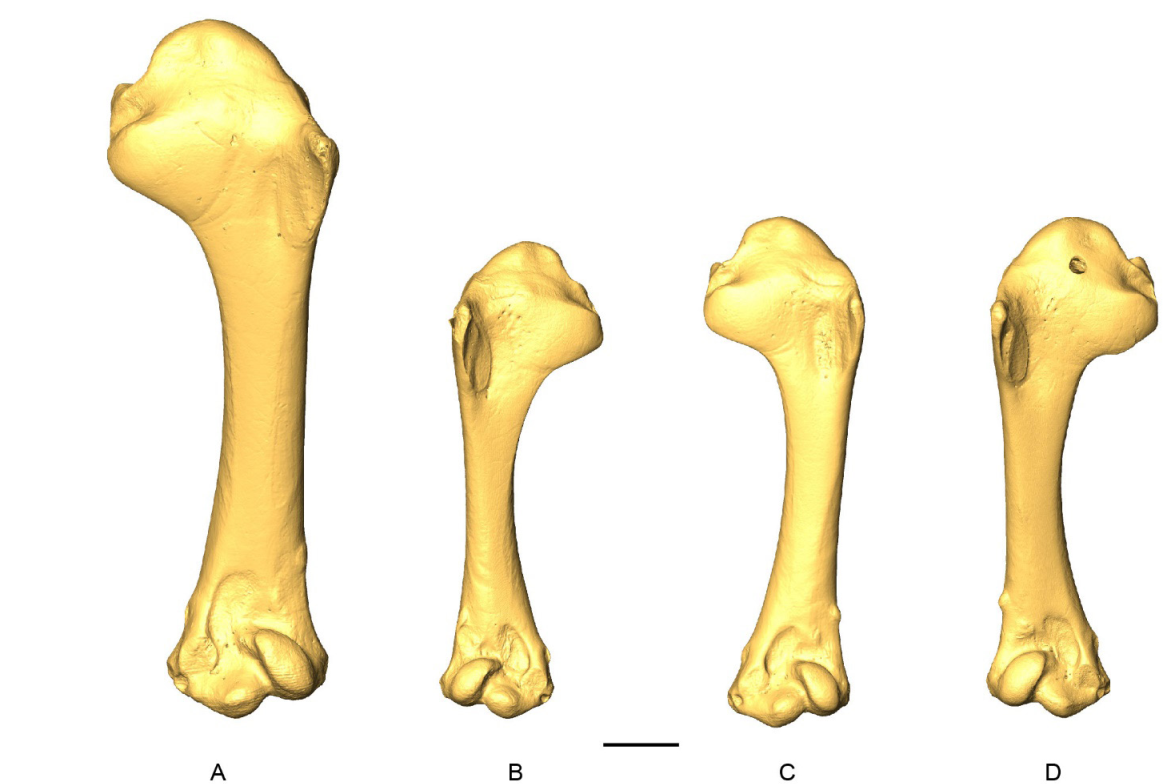
D-models of humeri in cranial view. A, Patagioenas inornata USNM 290977; B, Geotrygon montana MNHN-ZO-AC-1993-116; C, Leptotila wellsi NHMUK-ZOO-1898.9.20.1; D, Zenaida asiatica PACEA-O-707. B, D, right side; A, C, left side. Scale bar equals 5 mm.
APPENDIX K
Additional characters of humerus. The presence of an eminence on the ventral margin of crista bicipitalis is an additional proximal character of the humerus linked to HUM_VM observed in Zenaida asiatica that distinguishes it from Zenaida aurita and other 2 congeneric.
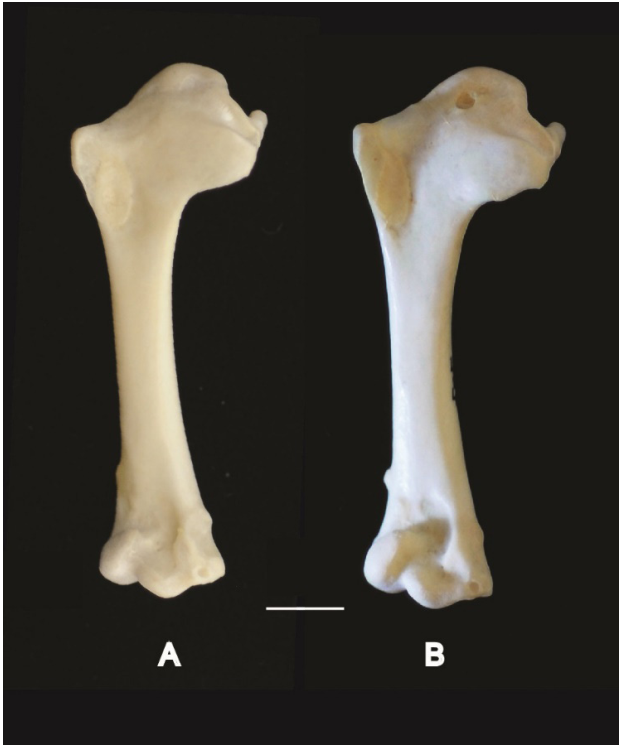
Right humeri in cranial view. A, Zenaida aurita PACEA-O-865; B, Zenaida asiatica PACEA-O-707. Scale bar equals 5 mm.
An additional distal character was recognized by Wetmore (1922). Based on the form of the fossa olecrani, this author suggested a shared behavior (flight) linking the three species of Patagioenas. The fossa olecrani in P. inornata bears an “inner wall merging with adjacent entepicondylar process in a gentle slope (forming a basin-like depression)” while in P. leucocephala and P. squamosa it has a “deeper, inner wall abrupt, forming a distinct pit at base of entepicondylar process” (Wetmore op. cit.: 318). These latter species are migratory unlike the P. inornata, which is more sedentary.
LITERATURE CITED
Wetmore, A. 1922. Bird remains from the caves of Porto Rico. Bulletin of the American Museum of Natural History, 46: 297-333.
Apendix L
APPENDIX M
3D-models of ulnae in cranial view. 3 A, Patagioenas inornata USNM 290977; B, Geotrygon chrysia USNM 292518; C, Leptotila wellsi NHMUK-ZOO-1898.9.20.1; D, Zenaida asiatica PACEA-O-707. B, right side; A, C-D, left side. Scale bar equals 5 mm.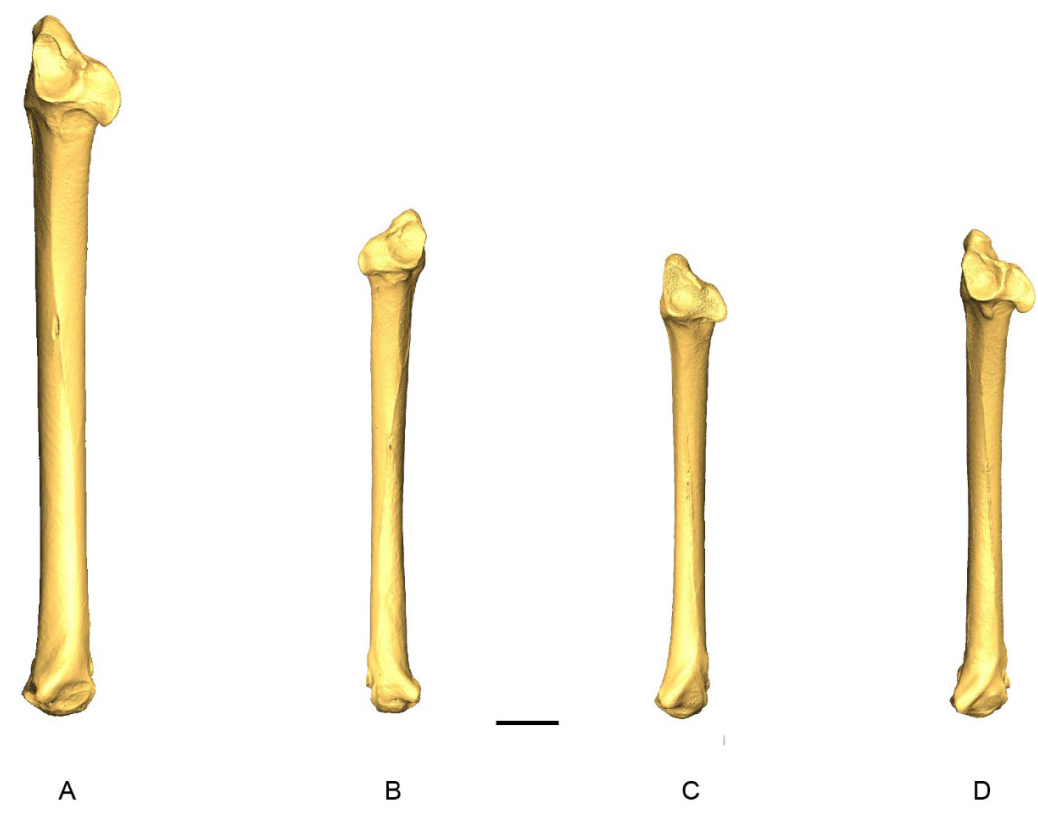
APPENDIX N
Measurements (in mm) and characters of carpometacarpus from modern specimens (character states see Fig. 15). Unavailable data is indicated by a dash, 3D-models with an asterisk
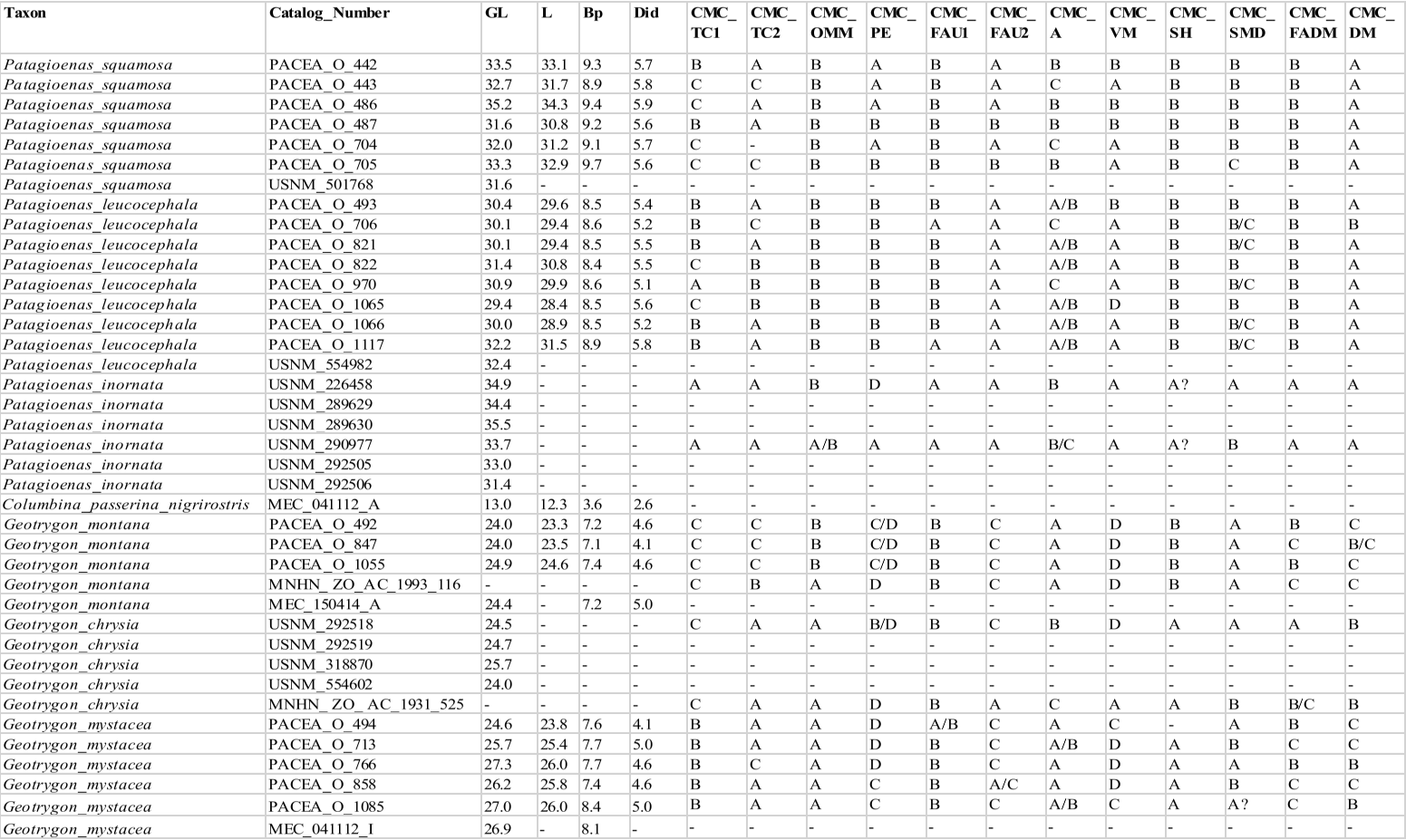
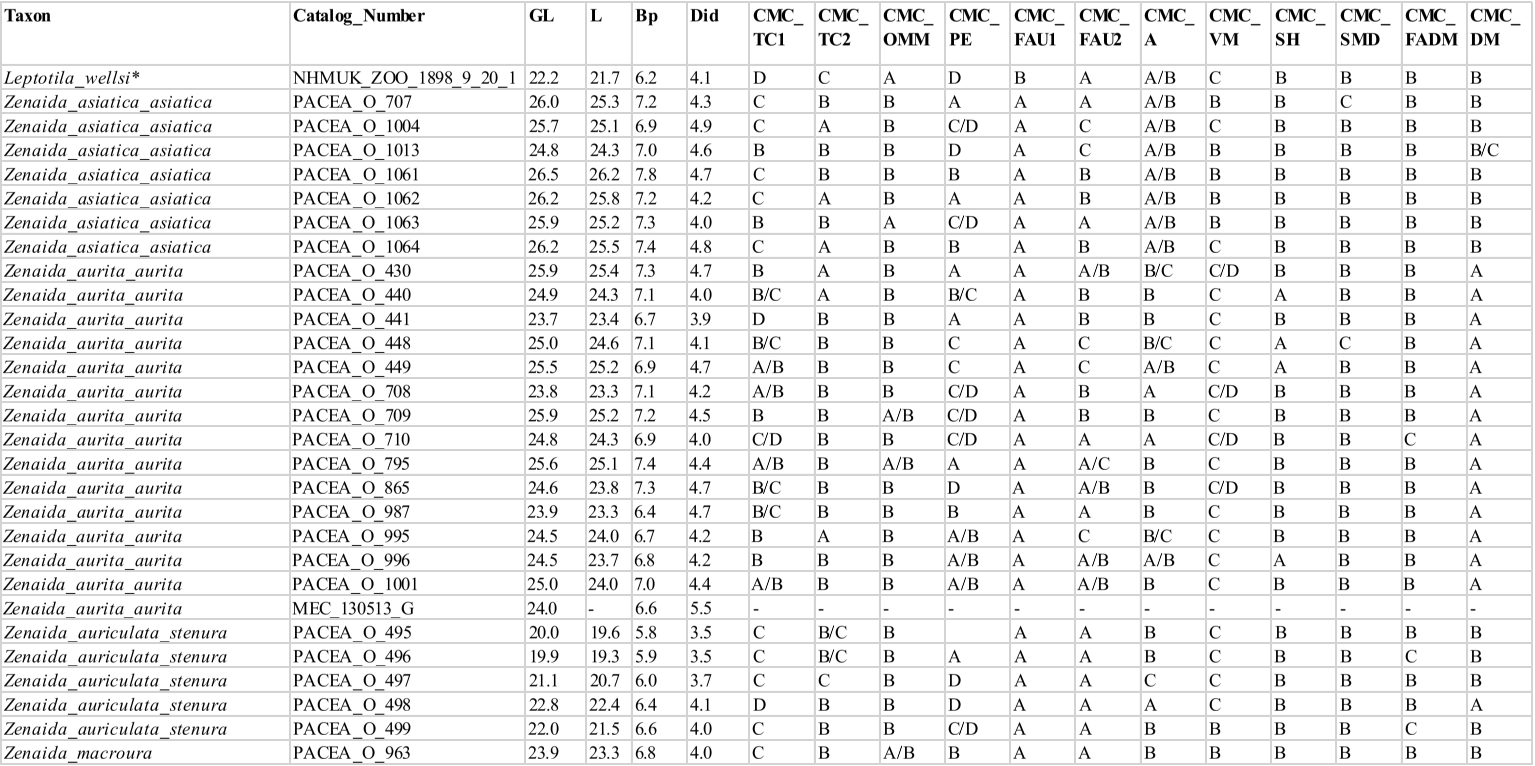
APPENDIX O
3 D-models of carpometacarpi in dorsal view. A, Patagioenas inornata USNM 226458; B, Geotrygon montana MNHN-ZO-AC-1993-116; C, Leptotila wellsi NHMUK-ZOO-1898.9.20.1; D, Zenaida asiatica PACEA-O-707. A-C, right side; D, left side. Scale bar equals 5 mm.
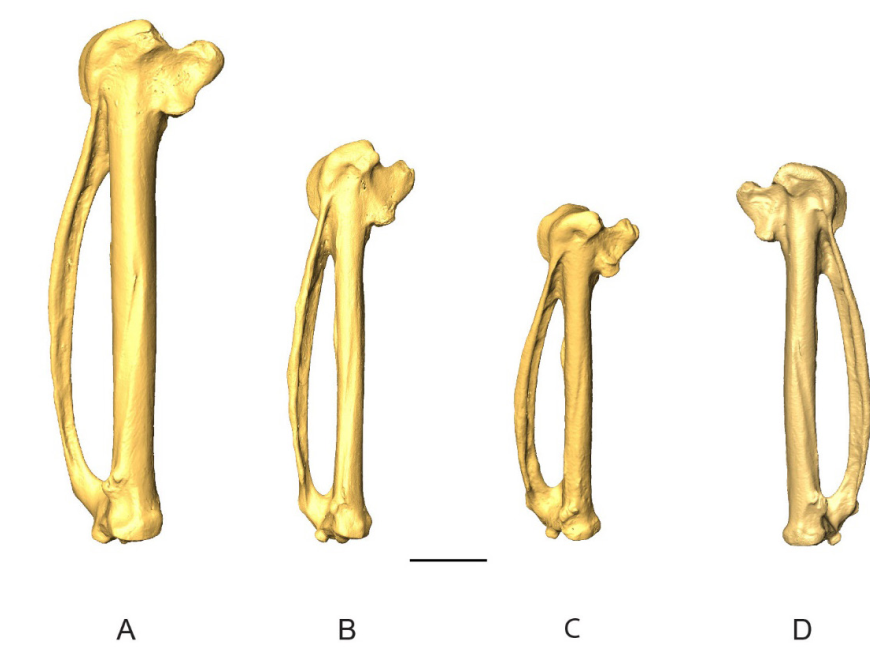
APPENDIX P
Measurements (in mm) and characters of femur from modern specimens (character states see Fig. 18). Unavailable data is indicated by a dash, 3D-models with an asterisk
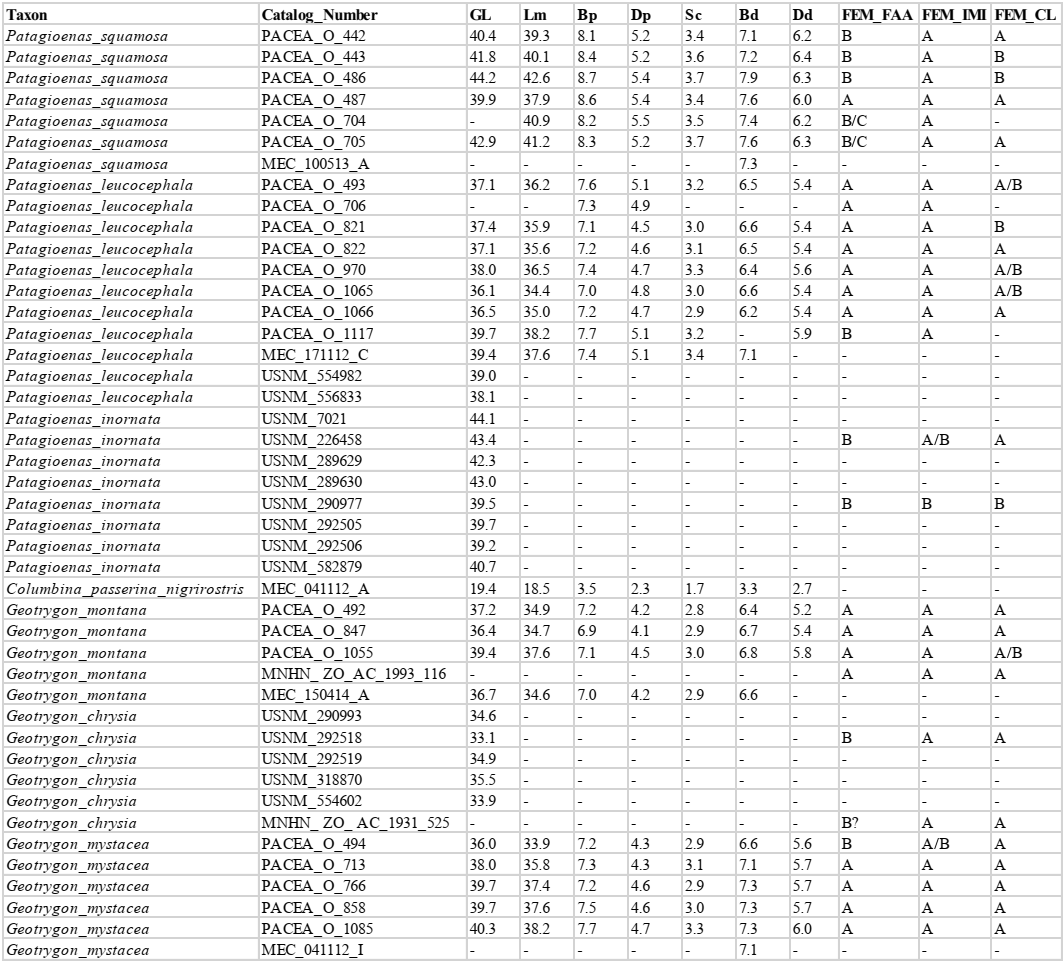
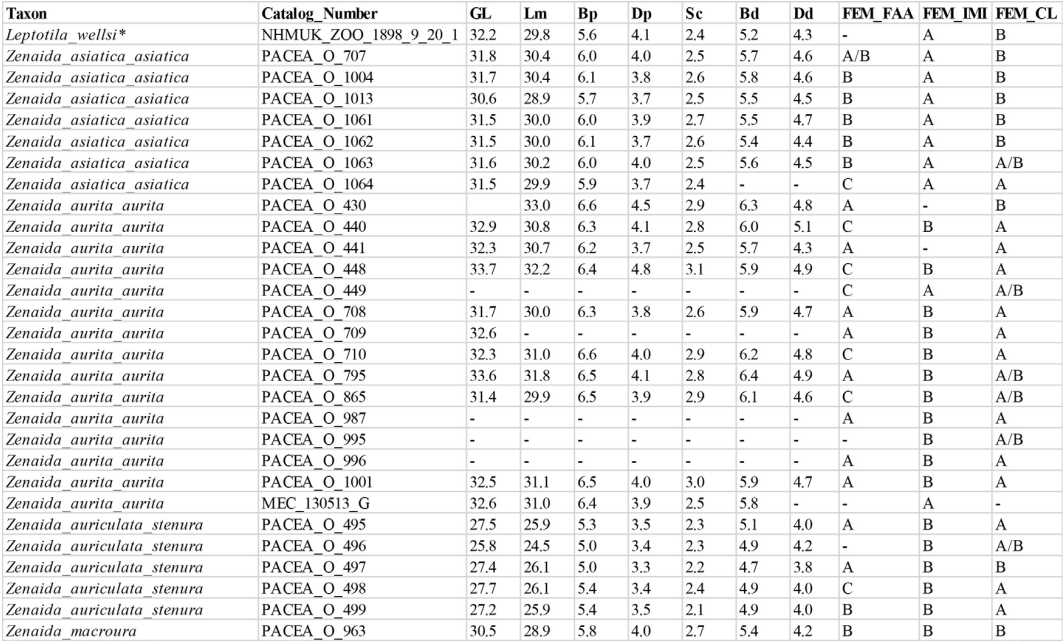
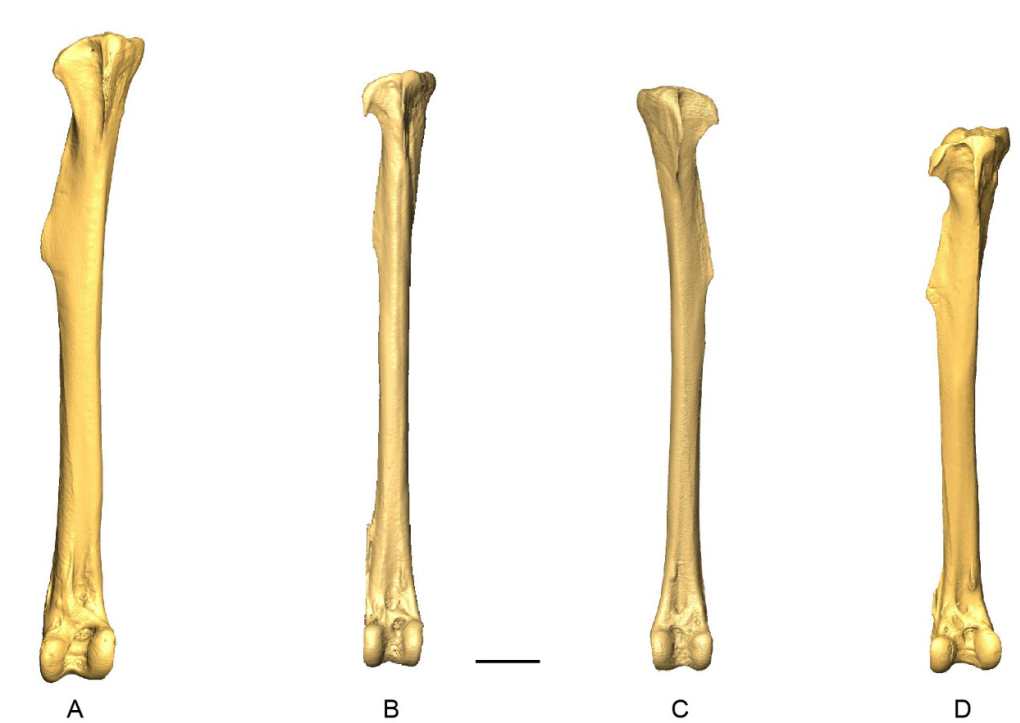
APPENDIX Q
3 D-models of femora in caudal view. A, Patagioenas inornata USNM 226458; B, Geotrygon montana MNHN-ZO-AC-1993-116; C, Leptotila wellsi NHMUK-ZOO-1898.9.20.1; D, Zenaida asiatica PACEA-O-707. A - B, D, right side; C, left side. Scale bar equals 5 mm.
APPENDIX R
Measurements (in mm) and characters of tibiotarsus from modern specimens (character states see Fig. 21). Unavailable data is indicated by a dash, 3D-models with an asterisk
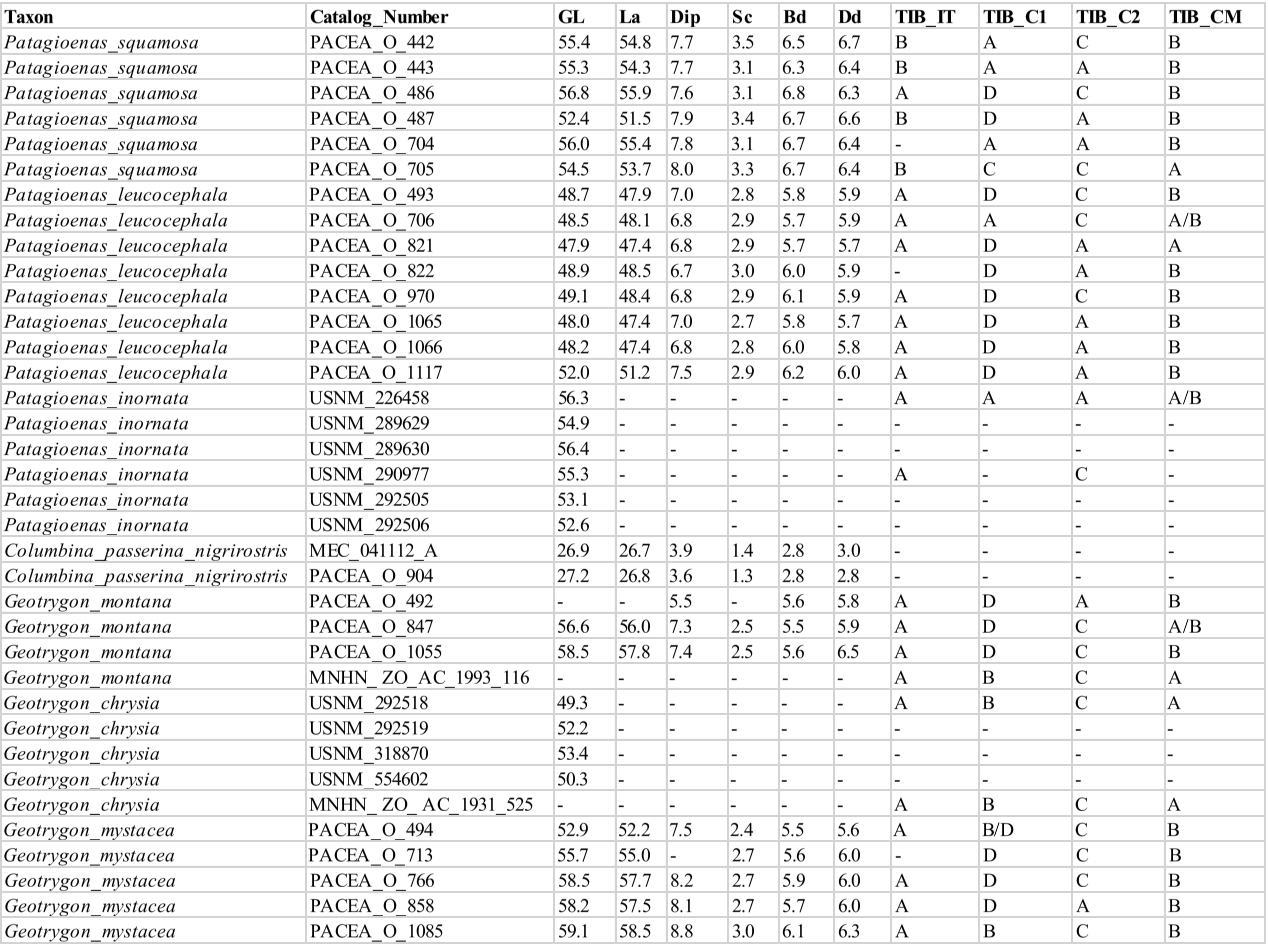
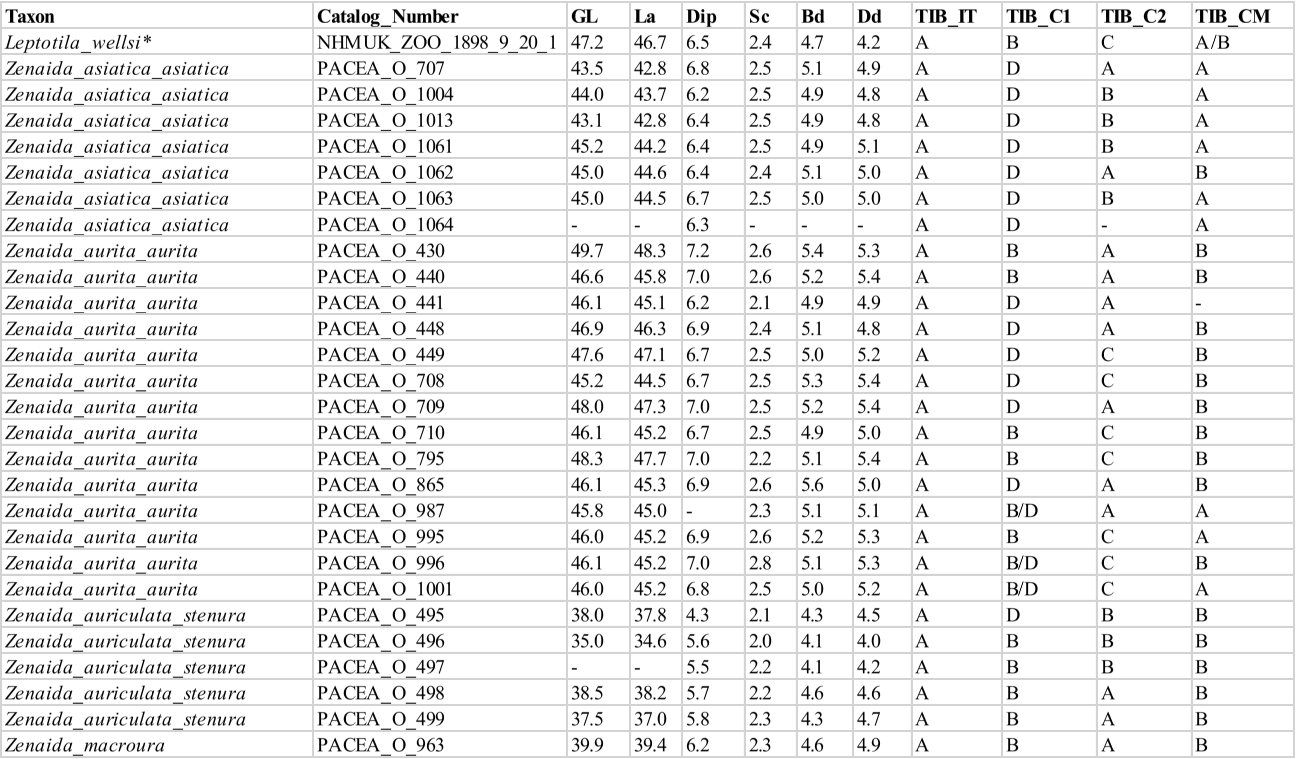
APPENDIX S
D-models of tibiotarsi in cranial view. 3 A, Patagioenas inornata USNM 226458; B, Geotrygon montana MNHN-ZO-AC-1993-116; C, Leptotila wellsi NHMUK-ZOO-1898.9.20.1; D, Zenaida asiatica PACEA-O-707. A - B, D, right side; C, left side. Scale bar equals 5 mm.
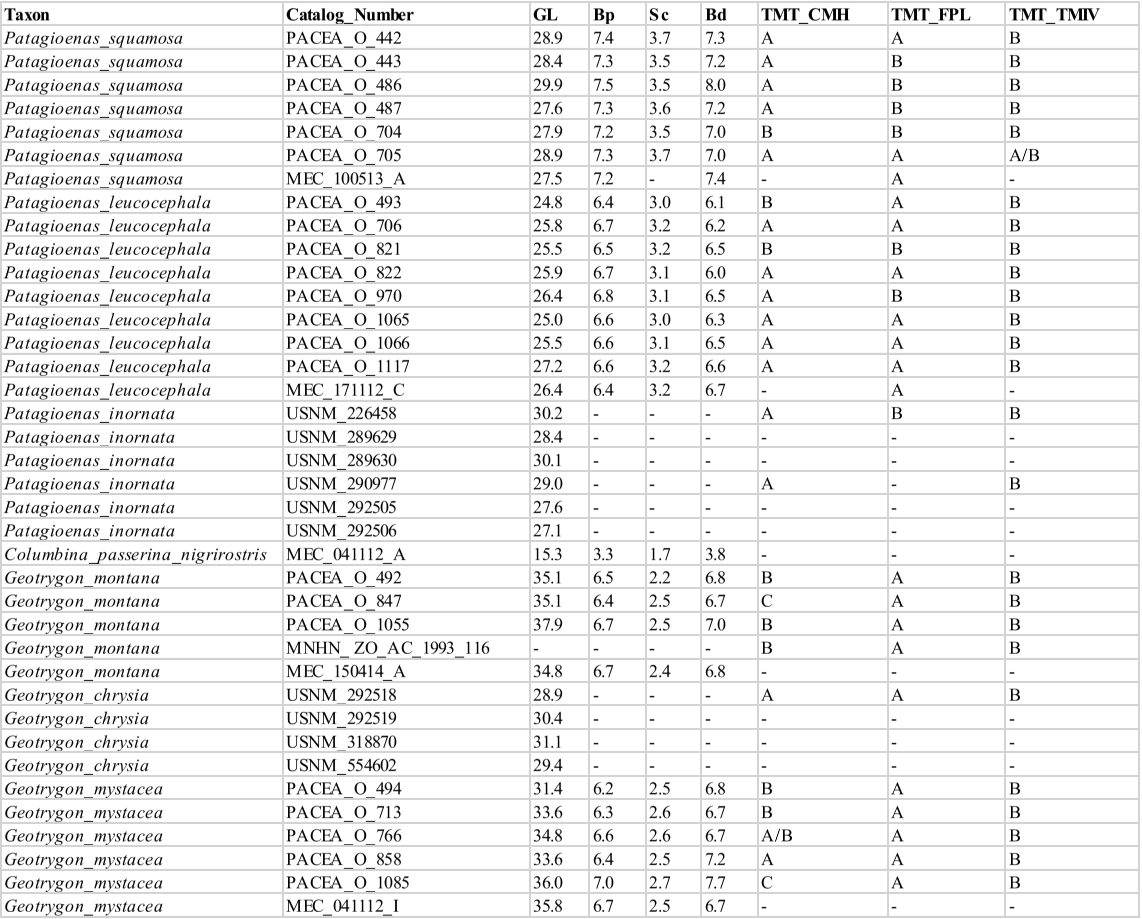
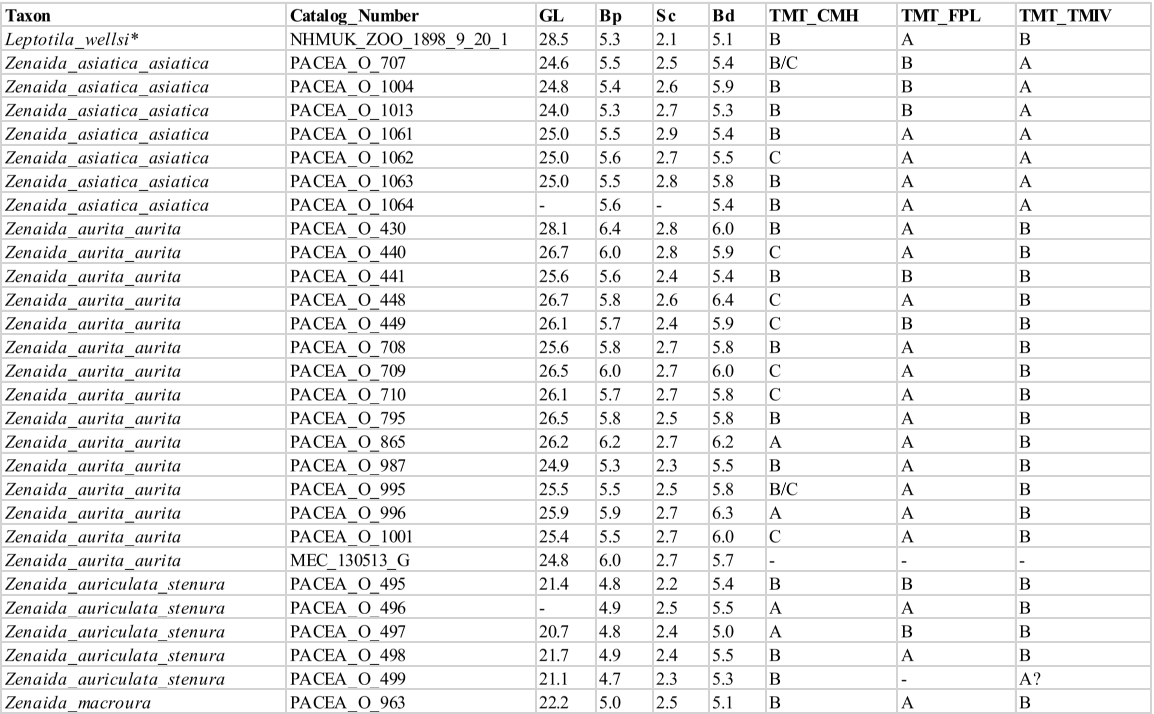
APPENDIX U
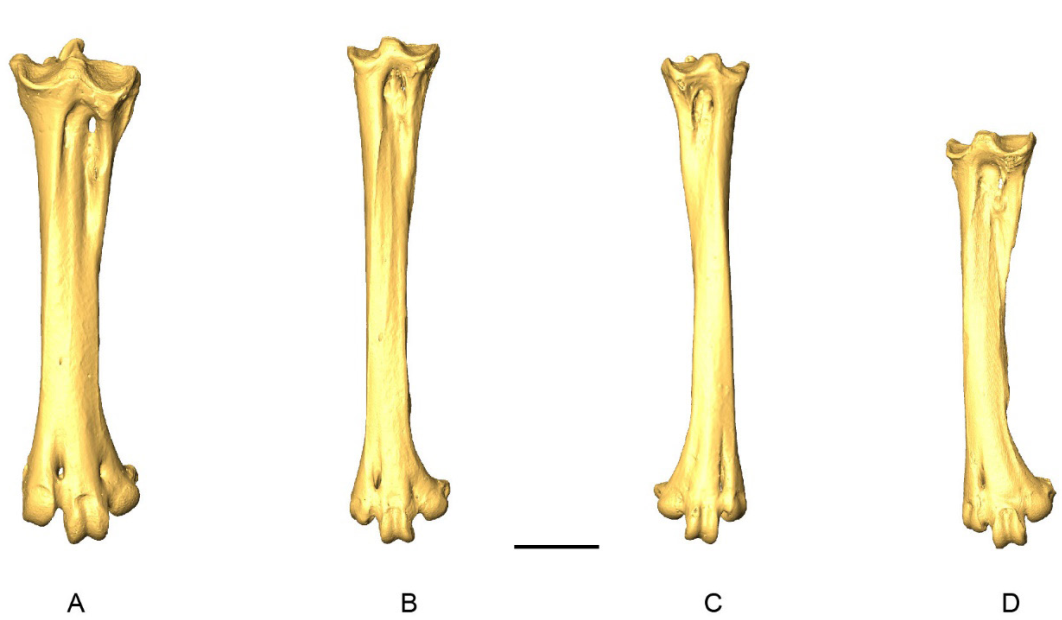 3
D-models of tarsometatarsi in
dorsal view.
A,
Patagioenas inornata
USNM 226458; B,
Geotrygon montana
MNHN-ZO-AC-1993-116; C,
Leptotila wellsi
NHMUK-ZOO-1898.9.20.1;
D,
Zenaida asiatica
PACEA-O-707.
A
-
B, D, right side; C, left side.
Scale bar equals 5 mm.
3
D-models of tarsometatarsi in
dorsal view.
A,
Patagioenas inornata
USNM 226458; B,
Geotrygon montana
MNHN-ZO-AC-1993-116; C,
Leptotila wellsi
NHMUK-ZOO-1898.9.20.1;
D,
Zenaida asiatica
PACEA-O-707.
A
-
B, D, right side; C, left side.
Scale bar equals 5 mm.