
INTRODUCCIÓN
Para Cuba, la lista de reptiles cuenta con 161 especies descritas (Torres López et al., 2017), de las que 141 son endémicos (88 %), ocho han sido introducidas por el hombre y 12 son nativas. Estos datos sitúan a los reptiles como el segundo grupo de vertebrados terrestres más diverso, solo superado por las aves; y el segundo en cuanto a endemismos, superado por los anfibios.
Los lagartos Anolis son el segundo género más diverso de vertebrados en el Caribe, superado solo por las ranas del género Eleutherodactylus. Además, el grupo constituye un caso ideal para estudios de biodiversidad debido a la alta densidad con que se manifiestan (Schoener, 1983; Reagan, 1992). Pianka (1989) plantea que la composición de la fauna de lagartos está profundamente influenciada por la estructura de la vegetación. Existen diversos estudios de relaciones ecológicas entre la variabilidad de las comunidades y la estructura de la vegetación (Krebs, 1985; Margalef, 1998). Según Arias (1997) las relaciones interespecíficas entre las poblaciones pueden explicar los mecanismos de regulación de las comunidades, pero generalmente no ofrecen explicación de los patrones de distribución espacial y temporal.
Rand (1964) comenzó los trabajos de subnicho estructural con comunidades de lagartos, definiéndolo como la posición exacta que ocupan las especies en el medio en cuanto a estrato, sustrato, altura y diámetro de las perchas. Posteriormente,teniendo en cuenta los recursos, se definieron cuatro subnichos: estructural o de microhábitat, trófico, climático y temporal (Pianka, 1973). En nuestro país, los estudios comunitarios de nicho comenzaron con el trabajo de Berovides y Sampedro (1980), en el que se analizan las cuatro formas de uso de los recursos por las especies de iguánidos de Cuba. Igualmente, sobresale el trabajo de Arias (2009), acerca de la relación de las especies de reptiles con cada tipo de formación vegetal en Cayo Santa María, así como el uso de los recursos y condiciones que utilizan las mismas.
Por otro lado, Losos (1990) planteó que los Anolis del Caribe se han diversificado para ocupar una amplia gama de microhábitats, que van desde el suelo del bosque hasta las copas de los árboles. Estas especializaciones de hábitat estaban descritas por Williams (1972) como “Ecomorfos”. Dicho autor define seis de estos ecomorfos y los denomina de la siguiente forma: Troncosuelo, tronco-dosel, tronco, gigantes de dosel (gigantes se refiere al tamaño de los lagartos), hierba-arbusto y ramitas. Losos et al., (2006) afirma que existe un patrón en el medio ecológico por el cual los miembros estrechamente relacionados del mismo tipo de ecomorfo coexisten.
La comunidad de lagartos de Loma de Cunagua presenta una gran variabilidad, tanto entre diferentes hábitats como dentro de uno mismo. En nuestro caso de estudio la variación de las comunidades de lagartos de Loma de Cunagua puede depender de las características de las especies que componen dicha comunidad o de las variaciones internas de los hábitats que condicionen la presencia y abundancia de las especies.
OBJETIVOS
Describir los patrones de distribución y abundancia de las especies de lagartos diurnos presentes en el Refugio de Fauna Loma de Cunagua.
Determinar cómo influyen la exposición solar y la disponibilidad de recursos en la segregación espacial de los lagartos diurnos en diferentes tipos de bosque del área de estudio.
Identificar los factores que resultan de importancia para la conservación de las especies y comunidades de lagartos diurnos en el área de estudio.
MATERIALES Y MÉTODOS
El Refugio de Fauna (RF) “Loma de Cunagua” se encuentra ubicado en la región central del país (Fig. 1), en el noreste de la provincia de Ciego de Ávila, en el municipio de Bolivia. El área posee una extensión de 8 228 ha terrestres. Constituye una zona núcleo dentro del Sitio Ramsar Gran Humedal del Norte de Ciego de Ávila yforma un domo salino de formaciones cársicas, que presenta una elevación máxima de 321 metros sobre el nivel medio del mar (Nerey-Contrera et al., 2014).
En el área existen tres formaciones vegetales: El bosque semideciduo sobre calizas, el bosque semideciduo sobre suelos de mal drenaje y los bosques de mangles. La primera constituye la formación vegetal más extendida y se presenta en dos variantes atendiendo a su grado de conservación: Bosque Semideciduo conservado y Bosque Semideciduo secundario. La densidad de este bosque es de baja a media y su altura oscila entre los 15-25 m. En dicha formación vegetal se destaca un estrato emergente compuesto por palmas y jocumas, así como un estrato arbóreo conformado entre otras, por las especies: ocuje (Callophyllum antillanum), yagruma (Cecropia peltata), guásima (Guazuma ulmifolia), cedro (Cedrelaodorata), majagua (Hibiscus elatus), almácigo (Bursera simaruba) y yaya (Oxandra lanceolata).
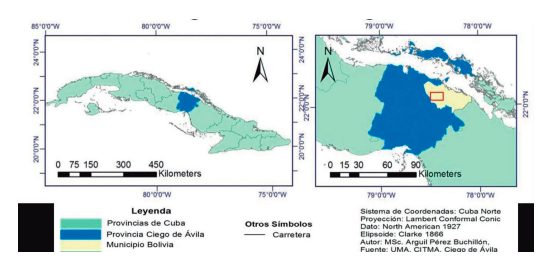
Figura 1. Ubicación geográfica del Refugio de Fauna Loma de Cunagua, como área de estudio de las variaciones espaciales de lagartos diurnos.
Las variantes de bosque semideciduo se diferencian principalmente en cuanto a la cobertura de dosel y la abundancia del estrato arbóreo, ambas características con marcado predominio en la variante conservada (Nerey-Contrera et al., 2014). Por otro lado, la variante secundaria presenta un mayor estrato herbáceo y un sotobosque más abierto, con menor densidad del sustrato rama, de lianas y epífitas. No obstante, ambas variantes presentan las mismas especies vegetales e indistintamente aparecen afloramientos cársicos, bastante abundantes en toda el área de la loma.
Para la clasificación sistemática de las especies de reptiles se siguieron los criterios de Rodríguez-Schettino (1999). Para la clasificación de las formaciones vegetales del área se consultó lo plateado en el plan de manejo antes mencionado, y se comprobó según los criterios de Capote y Berazain (1984). En cuanto a la diversidad de reptiles, el área cuenta con 18 especies, de los cuales 14 son endémicos de Cuba, para un 72.2 %. Destacando, dentro de los endémicos, las subespecies Anolis jubar cocoensis y Leiocephalus macropus hoplites, esta última con categoría de endémico local.
Para el estudio se marcaron 18 transectos de 200 metros de largo por cuatro metros de ancho en Bosque semideciduo sobre calizas, según lo establecido por Arias (2009). De ellos, 12 transectos en la variante conservada del Bosque semidecíduo, seis en la ladera Este y seis en la Oeste. En la variante secundaria del Bosque semidecíduo, solo se marcaron los seis de la vertiente Este, debido a la ausencia de dicha formación vegetal en la vertiente oeste de la loma. Los transectos se recorrieron desde las 9:00 a.m. hasta las 3:00 p.m., estandarizando el tiempo a 30 minutos. Los muestreos se realizaron con una frecuencia de dos visitas mensuales a cada transecto, extendiéndose durante un año, en el período entre mayo del 2015 hasta abril del 2016. En los recorridos se contó la cantidad de individuos de las distintas especies detectadas. Además, se tomó datos de hábitat, microhábitat y uso de los recursos, según lo establecido por Arias (2009).
Para determinar los parámetros cobertura de dosel, cobertura de suelo y cobertura de rocas, se utilizó un cilindro de 4.5 cm de diámetro, dividido en su extremo distal en cuatro cuadrantes. Las observaciones de estos parámetros se realizaron en los extremos de cada transecto, y además se agregó una medición extra en el centro de los mismos. Luego se promediaron los tres datos para obtener el valor a utilizar por cada transecto (Arias, 2009).
En la confección de las tablas de abundancia por especie se tomó el dato que respondía al mayor número de individuos en un transecto determinado. Para ello, se comparó los datos tomados en las dos visitas mensuales a cada transecto de ambas variantes de la formación vegetal. Se anotaron también los diferentes estados de las variables estrato y sustrato para el análisis del uso de los recursos estructurales (Arias, 2009). Para la variable estrato, se tomaron los estados árbol, arbusto y suelo; mientras que la variable sustrato se clasificó en los estados tronco, rama, hojarasca y piedra. Para determinar la actividad diaria en función del uso de los recursos, se tomó las alturas de las perchas en las que fueron observados cada uno de los individuos por especie detectados durante los conteos en cada uno de los 18 transectos, según lo establecido por Arias (2009). Las mediciones de altura se tomaron con una cinta métrica graduada en cm, desde el lugar exacto donde fue avistado el individuo hasta la superficie del suelo y de forma perpendicular. Con estas mediciones se confeccionaron tablas de alturas por especie, utilizando el promedio de altura para cada especie en ambas variantes de bosque.
La identificación de las tendencias principales de variación espacial de las comunidades de lagartos diurnos se realizó mediante la utilización de un análisis de ordenación (análisis de correspondencias AC), sobre la matriz de los transectos y la abundancia de las especies (Arias, 2009). Para la selección de las especies que caracterizan cada uno de los extremos de los ejes se consideraron los valores de sus contribuciones absolutas o inercias. Los resultados de los dos primeros ejes productos de la ordenación, han sido explicados en función de la estructura del microhábitat, mediante el análisis de correlación de Pearson (Sokal y Rohlf, 1979). La selección de este índice se debió a que todos los datos mostraron distribución normal al efectuar la prueba de Kolmogorov-Smirnov.
Para el análisis de la influencia climática sobre las especies de lagartos, solo se tuvo en cuenta la variable exposición solar. La misma se dividió en tres estados: Sol directo, Sol filtrado y Sombra. Dicha variable se tomó como factor determinante para la temperatura de los transectos y con ello, la temperatura en que se manifestaban los lagartos. Por su parte, en el análisis de uso de los recursos, se confeccionaron tablas de contingencia (Estados del recurso/Especie), y posteriormente, se aplicó la prueba “X1” para conocer si existieron diferencias significativas entre las especies de cada hábitat. El nivel de significación empleado fue del 5 %.
Para describir las variaciones de las especies entre los diferentes estados de los recursos y ver si existe competencia entre las mismas por su uso, se efectuó el cálculo de Amplitud de Nicho según la fórmula de Levins (1968). Los valores cercanos a 1 y a 0 indican un nicho espacial generalista y especialista respectivamente. Se toma como amplitud de nicho la variedad de estados de una variable a utilizar por cada especie. Se añadió también el cálculo de Solapamiento de Nicho de las especies, según la fórmula conocida como índice de solapamiento de Schoener (Krebs, 1999). Solapamiento de nicho se refiere al grado de coexistencia de varias especies en la utilización de un mismo estado de una determinada variable.
RESULTADOS
Descripción de la comunidad y su relación con la estructura de la vegetación. En el presente trabajo seregistraron para el Refugio de Fauna Loma de Cunagua 11 especies de lagartos diurnos, agrupadas en un orden, cuatro familias y cinco géneros. Es de destacar que no todas estas especies aparecen en la lista del Plan de Manejo del área protegida, por lo que representan nuevos reportes (Fundora-Caballero y Arias, inédito). El orden representado con la totalidad de las especies registradas fue Squamata. La familia de mayor representación fue Polychrotidae con nueve especies, seguida de las familias Tropiduridae y Teiidae con una especie cada una. El género más destacado fue Anolis con ocho especies.
El análisis de correspondencia utilizado para la ordenación de los transectos de muestreo en función de la composición de especies de lagartos diurnos muestra una fuerte absorción de varianza para los dos primeros ejes, con un 73.49 % (Fig. 2). El primer eje de ordenación recoge un 50.57 % y el segundo un 22.92 %, por lo que solo se consideraron estos ejes.
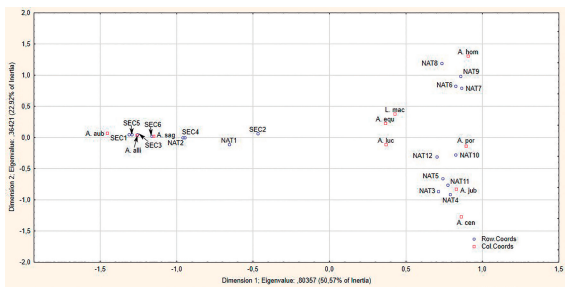
Figura 2. Representación de las coordenadas de las especies y las unidades de muestreo en los dos primeros ejes del Análisis de Correspondencias.
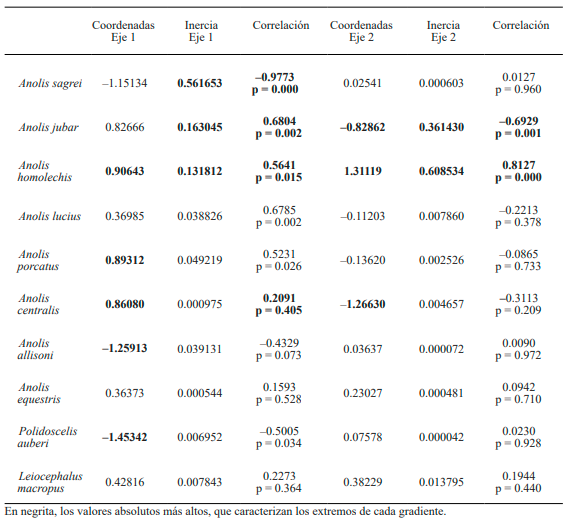
Al considerar el comportamiento de las especies que caracterizan los extremos del primer eje de ordenación, se observa una fuerte relación con el factor insolación. Especies como A. sagrei, A. allisoni y P. auberi aparecen en el extremo negativo del eje, prácticamente unidas a todos los transectos de la variante secundaria (Fig. 2). Cabe destacar que estas especies también aparecen asociadas a los dos primeros transectos de la variante conservada. Por otra parte, el extremo positivo del eje 1 está caracterizado por el resto de las especies, que aparecen muy cercanas a las unidades de muestreo de la variante conservada. El eje 2 por su parte muestra una relación entre las especies y la cobertura de dosel (Fig. 2). Así, aparecen separándose A. centralis y A. jubar de comunidades caracterizadas por A. lucius, A. equestris, A. sagrei, A. allisoni y P. auberi. A su vez, las mismas se separan de especies como A. homolechis y L. macropus que caracterizan el extremo positivo del eje 2 (Tabla I).
Tabla I. Coordenadas de las especies, inercia alcanzada y valores de correlación de Pearson en cada uno de los dos ejes principales del Análisis de Correspondencias
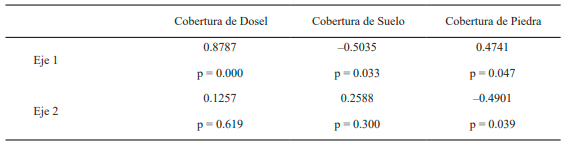
Por otro lado, existe una alta correlación entre las especies y las coberturas de dosel, suelo y piedra caracterizando el eje 1 del análisis de correspondencias (Tabla II). Para el eje 2, los valores de correlación solo son destacables entre las especies y la cobertura de piedra.
Tabla II. Valores de correlación de Pearson para las coberturas de dosel, suelo y piedras, en cada uno de los dos ejes principales del Análisis de Correspondencias
Segregación vertical de las especies de lagartos diurnos. En el presente estudio se tomaron datos de 1896 individuos de 10 especies (Tabla III). De estos, 1141 corresponden a la variante conservada y 755 a la variante secundaria, para un 60.18 % y 39.82 % respectivamente. Del total de especies, solo dos no pertenecieron al género Anolis. La variante conservada mostró la mayor cantidad de especies, por lo que unido al alto número de individuos encontrados sería el área a destacar para acciones de conservación. Por su parte, la variante secundaria presentó seis especies, de las cuales coinciden cuatro con la variante anterior.
Tabla III. Número de observaciones por especie y por cada variante de Bosque semideciduo de Loma de Cunagua, utilizados en los análisis espaciales
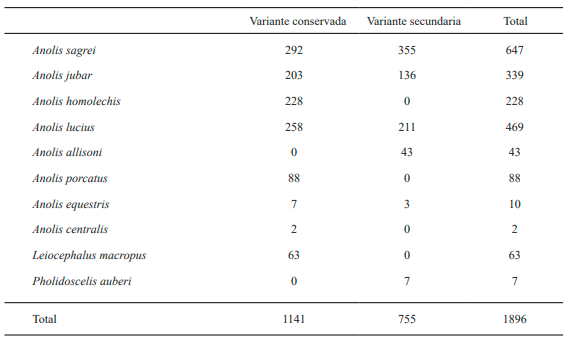
Se pueden diferenciar cuatro agrupaciones verticales de los lagartos diurnos presentes en la variante conservada del área de estudio, por las medias de las alturas a las que perchaban (Fig. 3). En primer lugar, se encuentra la especie L. macropus, que caracteriza la comunidad de lagartos a la altura cero. Los individuos de esta especie tienden a distribuirse entre el nivel de superficie del suelo y unos 0.40 metros por encima, casi siempre en presencia de rocas. A la anterior la acompañan individuos (en su mayoría juveniles) de la especie A. sagrei y muy escasos individuos de A. lucius.
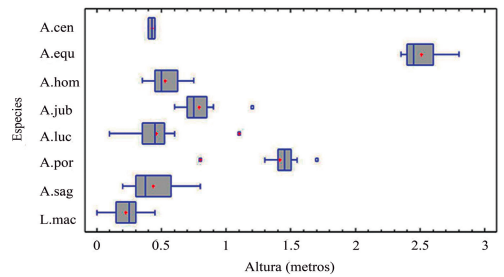
Figura 3. Representación de las alturas de las perchas registradas para cada especie en la variante conservada del bosque semideciduo de Loma de Cunagua. A. cen = Anolis centralis; A. equ = Anolis equestris; A. hom = Anolis homolechis; A. jub = Anolis jubar; A. luc = Anolis lucius; A. por = Anolis porcatus; A. sag = Anolis sagrei; L. mac = Leiocephalus macropus.
En segundo lugar, se encuentra la primera comunidad por encima de la altura cero hasta alcanzar una altura promedio que oscila entre 0.50 y 0.75 metros. La misma está caracterizada por A. sagrei, A. homolechis y A. lucius; acompañadas por A. centralis en menor cuantía, así como A. jubar que puede llegar hasta el metro de altura en sus perchas. Luego, viene una zona representada mayoritariamente por A. porcatusencontrados entre los 1.30 a los 1.50 metros. Por último, encontramos la comunidad por encima de los 2.0 metros de altura, caracterizada por la totalidad de los individuos de A. equestris, así como algunos ejemplares de A. porcatus.
Al analizar la Fig. 4, encontramos tres agrupaciones de alturas medias de los lagartos diurnos en la variante secundaria de Loma de Cunagua. Una primera que coincide con la variante anterior, a la altura cero y que estaría caracterizada por la especie P. auberi y muy escasos individuos, siempre juveniles, de A. sagrei y A. lucius.
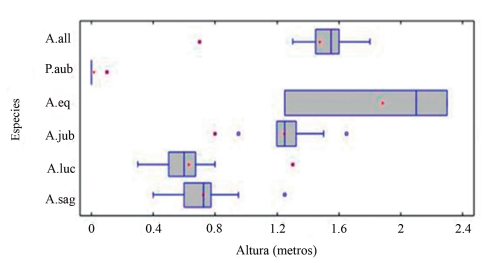
Figura 4. Representación de las alturas de las perchas registradas para cada especie en la variante secundaria del bosque semideciduo de Loma de Cunagua. A. all = Anolis allisoni; P. aub = Pholidoscelis auberi; A. eq = Anolis equestris; A. jub = Anolis jubar; A. luc = Anolis lucius; A. sag = Anolis sagrei.
En la segunda agrupación se aprecian diferencias con respecto a la variante conservada, puesto que, aunque se encuentra igualmente por encima de la altura cero, las comunidades de esta variante se encontrarían entre un 0.50 y 1.30 m de altura. No obstante, esta comunidad se caracterizaría, por las especies A. sagrei, A. lucius y A. jubar. En la tercera agrupación encontramos a una comunidad representada mayoritariamente por A. equestris y la generalidad de los individuos de A. allisoni. No obstante, esta agrupación presenta características ecotonales, debido a que pueden encontrarse individuos aislados de A. jubar. Asimismo, se observó que algunos ejemplares de la especie A. allisoni se movían también por perchas a las alturas cercanas a 1.40, lo que equivale a la segunda agrupación mencionada en esta variante.
Segregación y nichos espaciales de las especies de lagartos. Luego de analizar los resultados del cálculo de la amplitud de nicho, se obtuvo que las especies de menor amplitud en el uso del estrato son P. auberi y L. macropus. Este resultado coincide con lo encontrado en trabajo previopara las especies de suelo (Arias, 2009). De manera similar se comportaron también las especies de bajo número de muestra, como A. centralis y A. equestris (Tabla IV). Estos bajos valores de amplitud en el uso del estrato se observan para A. equestris en ambas variantes de la formación vegetal. Las restantes especies que presentaron la misma conducta solo se manifestaron en una sola de las variantes. En el mismo caso se encuentran A. porcatus y A. allisoni, que utilizan estratos similares, pero apareciendo solo en las variantes conservada y secundaria respectivamente. Así, A. porcatus tiende a comportarse como generalista y la segunda presenta un valor de uso de estrato que la podría situar como intermedia. Por su parte, A. homolechis también se comporta como generalista en el uso de los estratos, pero solo apareciendo en la variante conservada del área de estudio (Tabla IV).
Tabla IV. Valores de amplitud del uso de los recursos de las especies de lagartos, calculados en función de diferentes tipos de variables
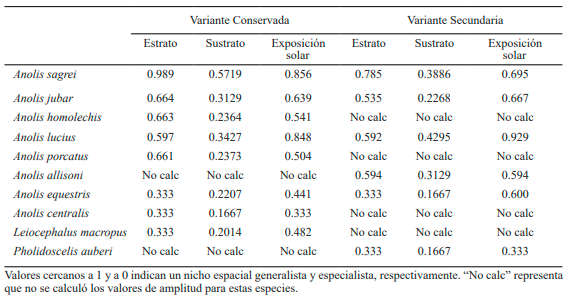
No obstante, en el caso de las especies que se avistaron en ambas variantes de bosque, A. sagrei y A. jubar presentan valores algo más altos en la variante conservada, aunque en ambas se manifiestan como generalistas para el uso del estrato. En el caso de A. lucius presenta valores de amplitud de uso del estrato casi iguales en ambas variantes. Esto, hace que se comporte como una especie intermedia entre un generalista y un especialista. Por otra parte, A. sagrei mostró valores que lo sitúan como especie intermedia en la amplitud del sustrato en la variante conservada.
No obstante, en la variante secundaria se muestra como especialista, manifestando un valor bastante bajo de amplitud del sustrato. Asimismo A. jubar y A. lucius se manifiestan en ambas variantes como especialistas (Tabla IV). De igual manera, se comportó A. equestris, aunque con número de muestras mucho menores como ya se dijo anteriormente.
Con respecto a la amplitud de sustrato de las especies A. homolechis, A. porcatus y A. centralis, las mismas presentaron valores bajos que los ubican como especialistas en la variante conservada. Un caso similar manifestó A. allisoni, pero en la variante secundaria. Por su parte, las especies de suelo como P. auberi y L. macropus mostraron valores bajos de amplitud en el uso del sustrato, solo superados por A. equestris y A. centralis. Esto, hace que se comporten como fuertes especialistas para la variable sustrato (Tabla IV). Cabe recordar que A. equestris y A. centralis aparecieron muy poco durante el tiempo de muestreo planificado, lo que podría influir en los análisis de amplitud de los recursos.
En cuanto a la amplitud en la condición exposición solar se observa que las especies con mayores valores en ambas variantes fueron A. lucius y A. sagrei, mostrándose como generalistas fuertes. Asimismo, se manifestó A. jubar con valores altos y casi iguales para ambas variantes de bosque (Tabla IV). En el caso de A. equestris mostró un valor mayor en la variante secundaria, lo que hace que en la misma se comporte como generalista. Para el caso de las especies que aparecieron en una sola de las variantes muestreadas, como A. homolechis, A. porcatus y L. macropus, las dos primeras muestran valores muy similares, que las sitúan como intermedias en la variante conservada. En el caso de L. macropus, presentó un valor relativamente bajo de amplitud a la exposición solar, por lo que podría comportarse como especialista. Asimismo, la especie A. centralis se muestra como especialista en la variante conservada, mientras que A. allisonilo hace como generalista en la variante secundaria.
Un análisis del solapamiento de estratos, para las diferentes especies de lagartos de la variante conservada, muestra que para A. sagrei se obtienen los mayores valores al comparar con A. lucius. De manera similar sucede, si lo comparamos con otras especies ecológicamente similares, como A. jubar y A. homolechis, así como con A. porcatus (Tabla V). Estas últimas especies también muestran altos valores de solapamiento entre ellas, destacando el valor entre A. homolechis y A. porcatus. Otras especies con valores altos de solapamiento en el uso del estrato serían A. equestris y A. jubar. Por otro lado, con valores más discretos, encontramos a las especies A. centralis, A. homolechis y A. porcatus. En el caso particular de L. macropus solo se distingue un valor alto de solapamiento al compararlo con A. lucius.
Tabla V. Valores del porcentaje de solapamiento espacial de los estratos entre especies de lagartos de la variante conservada de bosque semideciduo de Loma de Cunagua
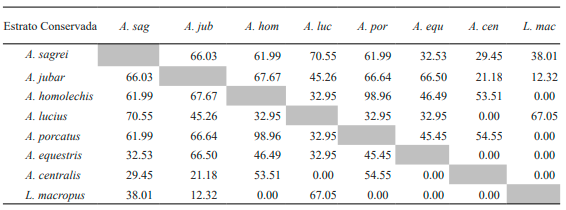
En la variante secundaria, el solapamiento del uso del estrato de A. sagrei es más alto con A. jubar, coincidiendo con la variante anterior. No obstante, A. sagrei también presenta valores moderadamente altos al compararlo con las especies A. lucius y A. allisoni (Tabla VI). Por su parte, A. jubar también muestra valores significativos si lo comparamos con las especies A. allisoni y A. equestris. Al igual que la especie de suelo de la variante anterior, P. auberi exhibe un solo valor a destacar en el solapamiento del uso del estrato, y es con la especie A. lucius.
Tabla VI. Valores del porcentaje de solapamiento espacial de los estratos entre especies de lagartos de la variante secundaria de bosque semideciduo de Loma de Cunagua
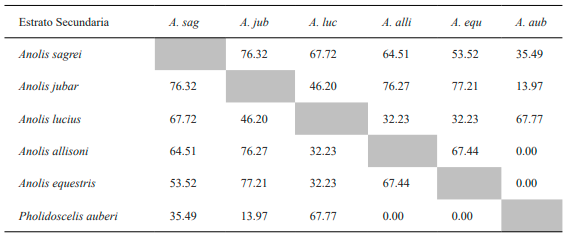
El cálculo del porcentaje de solapamiento para los sustratos de la variante conservada muestra que A. sagrei presenta su valor más alto con respecto a A. jubar (Tabla VII). La especie A. jubar también adquiere valores bastante altos al compararla con el resto de las especies que usa el estrato arbóreo o arbustivo, a excepción de A. lucius. De igual manera, ocurre con estas especies entre si y siempre excluyendo a A. lucius. Por último, L. macropus no muestra valores de importancia en el uso del sustrato con el resto de las especies presentes en esta variante.
Tabla VII. Valores del porcentaje de solapamiento espacial de los sustratos entre especies de lagartos de la variante conservada de bosque de Loma de Cunagua
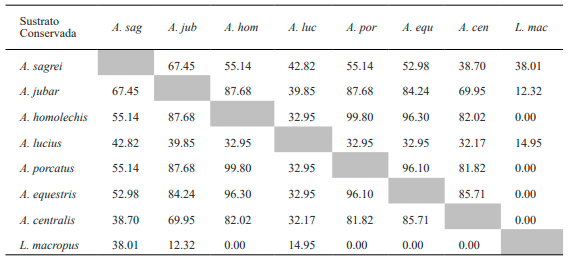
En lo referente al solapamiento del sustrato en la variante secundaria se observa que
A. sagrei vuelve a mostrar altos valores con las especies arborícolas, principalmente con A. jubar. En este caso, la relación es mayor incluso que lo observado para la variante anterior (Tabla VIII). Por su parte, A. jubar muestra el valor más alto de solapamiento en el uso del sustrato con A. equestris. Finalmente, tanto A. lucius como P. auberi no presentan altos valores al compararlas con el resto de las especies, lo que coincide con la variante conservada.
Tabla VIII. Valores del porcentaje de solapamiento espacial de los sustratos entre especies de lagartos de la variante secundaria de bosque de Loma de Cunagua
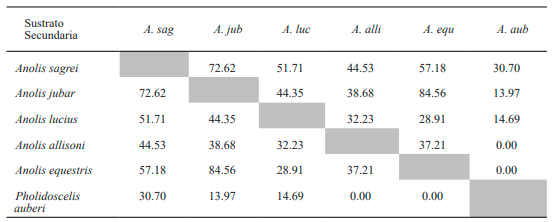
Al analizar el solapamiento en el uso de la exposición solar se obtuvo que A. sagrei presentó el más alto porcentaje al compararlo con A. lucius (Tabla IX). Igualmente, A. jubar mostró altos valores con respecto a las especies A. homolechis, A. porcatus, A. centralis y L. macropus. Además, en todas estas especies se observaron altos porcentajes de solapamiento en el tipo de insolación entre sí. Por otro lado, A. equestris no mostró valores significativos en la condición de insolación al comparase con el resto de las especies de la variante conservada.
Tabla IX. Valores de porcentaje de solapamiento espacial en el tipo de insolación entre especies de la variante conservada de bosque de Loma de Cunagua
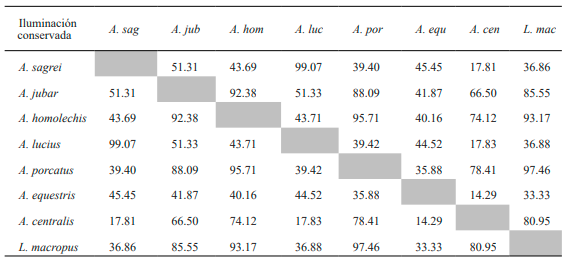
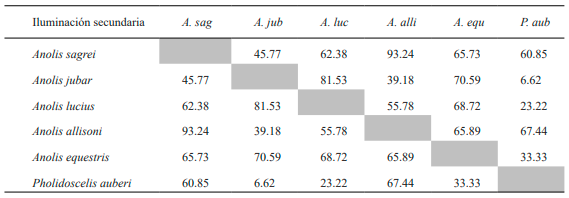
Con respecto a los resultados obtenidos en la variante secundaria para la condición insolación A. sagrei muestra, a diferencia de la variante anterior, elevados porcentajes con respecto a todas las especies exceptuando A. jubar. Por su parte, esta última solo muestra relación con A. lucius y A. equestris (Tabla X). En el caso de A. allisoni manifestó altos valores de solapamiento en el tipo de insolación con A. equestris y P. auberi.
Tabla X. Valores de porcentaje de solapamiento espacial en el tipo de insolación entre especies de la variante secundaria de bosque de Loma de Cunagua
DISCUSIÓN
El hecho de haberse encontrado un mayor número de especies en la variante conservada puede estar dado porque la misma presenta mayor desarrollo arbóreo y arbustivo. Por ello, brindaría a las especies de lagartos una mayor variabilidad de hábitat y microhábitat. Por otra parte, que existan cuatro especies de Anolis que coincidieron en ambas variantes de Bosque Semideciduo, puede deberse a que constituyen lagartos con ecomorfos similares. Además, estas especies se comportan mayormente como generalistas en cuanto a la tolerancia al factor insolación. Quizás, una excepción sería A. jubar, especie que solo se encontró en la variante secundaria hacia el interior de los transectos donde había una mayor cobertura de dosel, lo que filtraría más la exposición de la especie al sol. Esto, coincide con lo encontrado por Socarrás (1994) donde caracteriza a la subespecie A. j. cocoensis como umbrófila.
La ubicación de las especies en el Análisis de correspondencias (Fig. 2), muestra una clara relación con el factor insolación, lo que concuerda con lo encontrado por Arias (2009) para las comunidades de reptiles de Cayo Santa María. Así, las especies que caracterizan el extremo negativo son marcadamente heliófilas, ubicándose en la gráfica prácticamente solapadas con los transectos de la variante secundaria que presentaron menor cobertura de dosel. De igual forma, caracterizan el extremo positivo especies como A. jubar, A. porcatus, A. centralis y A. homolechis, relacionadas con los transectos de la variante conservada por comportarse claramente como umbrófilas.Además, en la misma figura se observan A. equestris, A. lucius y L. macropus,ubicadas relativamente cerca de las unidades de muestreo de mayor conservación, pero formando un claro grupo intermedio. Esto podría manifestar unaconducta generalista con respecto a la incidencia del sol, debido a coberturas de dosel intermedias que provocarían condiciones de sol filtrado. Arias (2009), encontró resultados similares de exposición solar para A. equestris y L. macropus. La relación existente entre los transectos de la variante natural con respecto a la cobertura de dosel está dada por el alto grado de conservación de esta variante. La cobertura de piedra se manifiesta en ambas variantes de bosque, debido a que el sustrato más generalizado de dicha formación vegetal está constituido por rocas calizas.
Aunque las especies A. sagrei y A. lucius se presentaron bajo las tres condiciones de insolación propuestas, prefirieron el sol directo, debido probablemente a su condición heliófila. Esto coincide con lo encontrado por Arias (2009) en Cayo Santa María. Por su parte, A. jubar, A. homolechis y A. porcatus se mostraron mayoritariamente en condiciones de sombra y sol filtrado, ya que se comportan como especies umbrófilas. Esto resultaría en una segregación por el factor iluminación, de especies ecológicamente similares como A. jubar, A. homolechis y A. sagrei. Otra especie umbrófila como L. macropus también prefirió las condiciones de sombra y de sol filtrado, respectivamente. Esto puede estar dado porque constituye una especie corredora, que al no estar elevada del suelo es más proclive al calentamiento que las especies que usan otros sustratos, escogiendo por ello los transectos con menor incidencia solar. Igualmente, A. equestris se mostró en sol filtrado, quizás debido a sus necesidades de termorregulación, en una variante donde existe mayor cobertura de dosel en las unidades de muestreo, y por consiguiente, mayores condiciones de sombra. Arias (2009), también mostró resultados similares con respecto al uso de la exposición solar de esta especie.
En la variante secundaria la especie A. sagrei prefirió los transectos con incidencia de sol directo y sol filtrado, debido a su amplia tolerancia a las altas temperaturas. Este resultado es coincidente con varios estudios, por ejemplo (Schwartz y Henderson, 1991; Arias, 1997, 2009), que tratan a la especie como heliófila. Por otro lado, A. lucius estaría utilizando más las condiciones de sol filtrado y sombra, debido quizás a la relación entre la cobertura de dosel y la de rocas, presente en los transectos de esta variante como ya se trató anteriormente. El hecho de que A. jubar se haya manifestado con mayor frecuencia en los transectos donde existía sol filtrado y sombra, puede estar dado porque la misma prefiera las áreas con mayor cobertura de dosel dentro de las unidades de muestreo de la variante secundaria. En los casos de A. allisoni y P. auberi se mostraron utilizando sol directo, así como sol filtrado en el caso de la segunda; porque ambas son especies heliófilas que dominaron los transectos de la variante secundaria. Ambos resultados concuerdan con Arias (2009) que clasifica a A. jubar como umbrófila y tanto a A. allisoni, como a P. auberi como marcadamente heliófilas.
Luego de analizar los resultados obtenidos en el cálculo de amplitud de nicho en el recurso estrato de ambas variantes de bosque, se sugiere que la presentación de menores valores por P. auberi y L. macropuspodría estar dada por su comportamiento como especialistas. Schwartz y Henderson (1991), ya clasificaban a estas especies como corredoras. En el caso de los bajos valores de amplitud de las especies A. equestris y A. centralis, podrían estar dados por la baja representatividad de las mismas, durante el tiempo de muestreo. Esto impediría resultados más concluyentes al comparar su amplitud en el uso de los diferentes recursos. Los altos valores obtenidos para A. homolechis y A. porcatus, podrían deberse a que manifestaron conductas generalistas en el uso de los diferentes estratos, pero limitándose a la variante conservada, debido a su condición umbrófila. Contrario a esto, los valores intermedios en la amplitud de A. allisoni para los diferentes estratos de la variante secundaria, podría deberse a que la especie manifestaría un comportamiento ecotonal, utilizando los estratos ocupados por su congénere A. porcatus en la variante anterior. No obstante, A. allisoni no apareció ni en suelo ni en perchas cercanas a dicho estrato, donde sí se mostró A. porcatus, pero en los transectos de vegetación conservada. Esto se puede explicar con lo planteado por Schoener (1977), quien demostró que el desplazamiento de los anólidos dentro de un hábitat, depende tanto de la estructura del propio hábitat como de la presencia o ausencia de competidores.
Los valores más altos en el cálculo de la amplitud del recurso estrato se mostraron para las especies A. sagrei y A. jubar, debido a que las mismas pueden considerarse como generalistas en el uso de estos recursos. Además, la alta disponibilidad del estrato arbustivo en los transectos de ambas variantes, hace posible que pese a ser competidores por hábitat, estas especies puedan segregarse en los tres estratos estudiados en función de otras variables como la cobertura de dosel.
Lo anterior, concuerda con los resultados de Losos (2004) que planteó la segregación de especies similares mediante la competencia por los recursos. En el caso particular de A. lucius, mostró valores intermedios de amplitud, ya que, a pesar de encontrarse en ambas variantes de bosque, usó preferiblemente el recurso suelo. Esto con excepción de los machos, que tienden a preferir el estrato arbóreo, pero casi nunca se encontraban sobre arbustos, a pesar de su gran disponibilidad de este recurso en la variante conservada.
En el cálculo de amplitud para el recurso sustrato la especie A. sagrei mostró valores medios, lo que probablemente se deba a que la misma prefirió los troncos y la hojarasca en la variante conservada. Esto último debido a la gran cantidad de juveniles encontrados, que muestran preferencia por dicho sustrato, probablemente para separarse de los adultos de su propia especie que representarían competencia para los recursos alimentarios. En el caso de la variante secundaria A. sagrei se comporta como especialista, mostrando la generalidad de sus individuos sobre los troncos, con escasas apariciones en ramas y hojarasca. Esto puede estar influenciado por el bajo número de juveniles y la alta aparición de machos, encontrados con respecto a la variante anterior. Dichos resultados coinciden con los de Estrada y Novo (1986) y Arias (2009), que trataron esta preferencia por los troncos de la especie A. sagrei. Por otra parte, que A. jubar y A. lucius presentaran bajos valores del índice de amplitud, indica que ambos se comportaron como especialistas. Así, el primer caso prefirió el sustrato tronco y el segundo, la presencia de piedras en los transectos de ambas variantes. Asimismo, A. equestris se manifestó mayoritariamente en troncos, pese a ser una especie de dosel. Esto podría estar influenciado por su baja representatividad durante el tiempo de muestreo. No obstante, Arias (2009) también referenció la preferencia de esta especie, tanto por troncos como por ramas.
También como especialistas se comportaron las especies A. homolechis, A. porcatus y A centralis, que mostraron bajos valores de amplitud, dado que prefirieron el uso del sustrato tronco. Cabe destacar que las tres especies solo aparecieron en la variante conservada, donde al parecer desplazaron en el uso de este sustrato a especies como A. sagrei y A. jubar. A. allisonise mostró como especialista, ocupando los troncos de la variante secundaria, debido a que A. porcatus no se presentó en esta variante. Esto coincide con los resultados de Losos (2004), debido a que tanto A. allisoni, como A. porcatus, constituyen especies ecológicamente similares que competirían con más fuerza por este recurso.
Igual que en el uso del estrato, las especies A. auberi y L. macropus se manifestaron como fuertes especialistas, dado sus bajos valores de amplitud en el uso del sustrato. Ello se debe a que ambas constituyen especies corredoras que solo utilizaron sustratos determinados y en variantes diferentes de bosque semideciduo. En el primer caso la especie prefirió el suelo desnudo de la variante secundaria, lo que aparte de espacio para desplazarse, le brindó el grado de insolación adecuado. En el caso de L. macropus, prefirió la hojarasca o las piedras, siempre en la variante conservada. Esto, probablemente debido a que la especie busque separarse más del suelo, evitando las altas temperaturas, y se manifieste entonces relacionada a la alta cobertura de piedras existente en los transectos de la variante conservada. El hecho de que las especies de mayor amplitud en cuanto a la condición exposición solar en ambas variantes de Bosque semideciduo fueran A. sagrei y A. lucius, debe estar dado porque constituyen las especies de mayor plasticidad en cuanto al factor insolación. Además, todas las especies presentaron valores altos en este aspecto, debido a que las estrategias de termorregulación de estos lagartos se basan en los cambios de exposición solar, como bien indica Arias (2009).
Los altos valores de solapamiento de estratos para la variante conservada de A. sagrei y A. lucius pueden estar dados por compartir mayoritariamente los estratos arbóreo y suelo, con densidades bastante semejantes entre ambas especies. De igual manera ocurrió entre A. sagrei con respecto a A. jubar y A. homolechis, debido a que utilizan los estratos estudiados de forma muy semejante. Resultado muy similar al encontrado por Losos (2004) y Arias (2009), que sitúan a estas especies como fuertes competidores por el hábitat. El hecho de que A. porcatus comparte altos valores de solapamiento con las especies antes mencionadas, está dado porque utiliza, aunque con menores densidades, los estratos arbóreos y arbustivo de la variante conservada. Además, es de resaltar que el 55 % de sus individuos constituyeron machos, al igual que el 45 % de las especies A. jubar y A. homolechis, así como el 50 % de los representantes de A. lucius. Esto coincide con lo planteado por Losos (2004), quien muestra que el solapamiento territorial entre machos es fuertemente convergente dentro de los ecomorfos de Anolis. Esto probaría, que además del complejo conjunto de rasgos morfológicos, ecológicos y de comportamiento que han evolucionado en estas especies; laestructura social también se desarrolla en conjunto con el uso de su hábitat.
El alto valor observado en el solapamiento del estrato entre A. equestris y A. jubar puede deberse a que ambas constituyen especies casi netamente arborícolas. En lo referente al elevado índice de A. lucius con respecto a L. macropus, puede estar dado porque ambas son reconocidas especies de suelo. Cabe resaltar que en este caso el solapamiento no fue de un 100 %, probablemente debido al uso frecuente del estrato arbóreo por los machos de A. lucius.
En el caso de la variante secundaria, coincide el alto valor de solapamiento entre A. sagrei y A. jubar, igualmente por su uso de los árboles y los arbustos, casi con iguales valores entre ambos estratos. Los valores ligeramente altos de solapamiento entre A. sagrei y las especies A. allisoni y A. lucius pueden deberse a que las tres comparten el uso casi exclusivo del estrato arbóreo en dicha variante. Lo anterior concuerda con lo encontrado por Arias (2009) para el Matorral Xeromorfo sobre Arena, donde existieron altos valores de solapamiento para la generalidad de las especies, en el uso del estrato de esta formación vegetal. Por su parte, el grupo A. jubar, A. allisoni y A. equestris también mostraron valores relativamente altos de solapamiento de estratos, debido a su marcado uso de los árboles. En lo que respecta a las especies A. lucius y P. auberi estas mostraron un alto valor de solapamiento por su amplio uso del recurso suelo, tal y como sucedió con L. macropus en la variante anterior.
El cálculo de los porcentajes de solapamiento para los sustratos de la especie A. sagrei, adquiere un valor alto con respecto a A. jubar, debido probablemente al uso de ambos de los mismos sustratos, con cifras semejantes. El grupo de A. jubar, A. homolechis, A. porcatus y A. centralis presenta también altos valores de solapamiento por el uso de los sustratos tronco y ramas por dichas especies. Esto los hace competidores fuertes por el recurso sustrato, pese a que los dos primeros y el último presenten diferentes clasificaciones en cuanto a sus ecomorfos. Buthler y Losos (2002) documentaron que los ecomorfos varían el grado de utilización de algunos recursos y que estas variaciones ocurren solo en algunos hábitats, dado que los mismos ecomorfos varían en su grado de territorialidad.
Con relación al uso de la condición exposición solar, el hecho de que el valor más alto de la variante conservada haya sido entre A. sagrei y A. lucius debe estar dado porque ambas especies utilizaron las tres condiciones de iluminación estudiadas de manera muy similar. El resto de los valores de solapamiento de ambas fueron bastante bajos, debido a que tanto A. sagrei como A. lucius mostraron altos valores en el uso del resto de los recursos con las demás especies de la variante. Además, el factor insolación sería determinante para disminuir las interacciones de estas dos especies con el resto. Los altos porcentajes del grupo A. jubar, A. homolechis,
A. centralis y L. macropus se deben probablemente al uso por estas especies, de las condiciones de sombra, y con menos preferencia de sol filtrado. Esto coincide con Arias (2009) que trata varias de estas especies como marcadamente umbrófilas. Por otro lado, el que A. equestris no haya manifestado altos porcentajes de solapamiento con el resto de las especies, puede deberse a que se comporta como generalista en el uso de la condición exposición solar y mostraría bajas interacciones con el resto de la comunidad. Cabe destacar que su baja densidad en los muestreos podría influir en los bajos valores de solapamiento.
Contrario a la variante anterior, A. sagrei mostró altos valores de solapamiento con todas las especies de la variante secundaria, excepto con A. jubar. Esto puede deberse a la tolerancia al factor insolación que presentan todas estas especies y que no muestra A. jubar, lo que sería un factor a destacar a la hora de la segregación en dicha variante. Este resultado es semejante al encontrado por Losos (2004) que explica la influencia de la competencia en la segregación de las especies semejantes de una comunidad. Los altos porcentajes de solapamiento de la especie anterior con A. lucius pueden deberse a que esta última se comporta como generalista en el uso del factor iluminación en la variante secundaria. Por otro lado, el elevado valor que muestran A. jubar y A. equestris estaría dado porque ambas especies se manifestaron durante el tiempo de muestreo, utilizando con mayor frecuencia la condición Sol filtrado.
La relación entre A. allisoni y P. auberi en cuanto al factor iluminación puede estar dada por las marcadas características heliófilas de ambas especies, que además solo se encontraron en la variante secundaria. Por otro lado, los bajos valores de solapamiento en la condición insolación entre P. auberi y las especies A. equestris y A. jubar, podrían deberse a que mientras la primera constituye una especie estrictamente heliófila, la segunda prefirió utilizar unidades de muestreo con sol filtrado. En el caso de A. jubar, es marcadamente umbrófila, y por ello mostraría el menor valor de solapamiento de esta variante al asociarla con P. auberi.
CONCLUSIONES
El grado de insolación del microhábitat, regulado por la cobertura de dosel, constituye el factor principal que afecta la organización de la estructura de las comunidades de lagartos diurnos de Loma de Cunagua. Existe una clara diferenciación en el uso de los recursos estrato y sustrato por las especies, caracterizando el suelo P. auberi, L. macropus y A. lucius, aunque este último también se manifiesta usando estratos vegetales como el resto de las especies. La altura y la segregación en el uso de los recursos constituyeron factores claves para la coexistencia y abundancia de las comunidades de lagartos diurnos dentro de los estratos de ambas variantes de bosque. El análisis del uso de los recursos y la exposición solar nos muestra la existencia de variaciones espaciales que posibilitan la coexistencia de las especies señaladas en el estudio, la disminución de sus interacciones y permite identificar los factores importantes para la conservación de la comunidad de lagartos diurnos del área.
AGRADECIMIENTOS
Se agradece a la Empresa Nacional de Protección para la Flora y la Fauna de Ciego de Ávila por permitirnos el acceso al área, así como a los especialistas de la Subdelegación de Medio Ambiente del CITMA de la misma provincia, por las pertinentes revisiones de este manuscrito. También quisiera agradecer a Javier Torres López (Universidad de Kansas), por su oportuna ayuda en aclaraciones finales de documento.
Literatura Citada
Arias, A. 1997. Ecología de las comunidades de lagartos del bosque siempreverde micrófilo de Cayo Santa María. Tesis de Licenciatura. Facultad de Ciencias Naturales y Matemática. Universidad de Oriente, Cuba, 34 pp.
Arias, A. 2009. Ecología de las comunidades de lagartos de Cayo Santa María, Villa Clara, Cuba. Tesis de doctorado. Universidad de Alicante y Universidad de Pinar del Río, 181 pp.
Berovides, Á. V. y A. M. Sampedro. 1980. Competición en especies de lagartos iguánidos de Cuba. Ciencias Biológicas, 5: 115-122.
Butler, M. A. y J. B. Losos. 2002. Multivariate sexual dimorphism, sexual selection, and adaptation in Greater Antillean Anolis lizards. Ecological Monography, 72: 541-559.
Capote, R. P. y R. Berazaín.1984. Clasificación de las formaciones vegetales de Cuba. Revista del Jardín Botánico Nacional, 5: 27-75.
Estrada, A. y J. Novo. 1986. Subnicho estructural de Anolis sagrei en Cayo Inés de Soto, Cuba. Análisis intra y extrapoblacional. Poeyana, 320: 1-13.
Krebs, C. J. 1985. Ecology. The experimental analysis of distribution and abundance. 3rd.ed. Harper and Row, Publisher, NY, 645 pp.
Krebs, C. J. 1999. Ecological methodology. 2nd.ed. Addison-Wesley Educational Publishers, Inc. Benjamin/Cummings. CA. pp. 410-454.
Levins, R. 1968. Evolution in changing environments. Princeton University Press. Princeton, N.J.132 pp.
Losos, J. B. 1990. Ecomorphology, performance capability, and scaling of West Indian Anolis lizards: an evolutionary analysis. Ecological Monography, 60: 369-388.
Losos, J. B. 2004. Adaptation and speciation in Caribbean Anolis lizards (pp 335-343). En: Dieckmann, U., M. Doebeli, J. Metz, & D. Tautz (Eds.), Adaptive speciation. Cambridge University Press.
Losos, J. B., R. E. Glor, J. J. Kolbe, y K. Nicholson. 2006. Adaptation, speciation, and convergence: a hierarchical analysis of adaptive radiation in Caribbean Anolis lizards. Annual Monography Botanical Gardens, 93: 24-33.
Margalef, R. 1998. Ecología. 9na. Reimpresión. Ediciones Omega, Barcelona, 951 pp.
Nerey-Contrera, W., J. H. Irarragorri, F. G. Quiñones, I. R. Companioni, O. L. Carvajal,
J. L. Moreno, O. M. Suárez, y O. B. Pupo. 2014. Plan de Manejo Refugio de Fauna “Loma de Cunagua”. Empresa Nacional para la Protección de la Flora y la Fauna. Ciego de Ávila. 2014-2018.
Pianka, E. R. 1973. The structure of lizard communities. Annual Review Ecological Systems, 453-474.
Pianka, E. R. 1989. Desert lizard diversity: additional comments and some data. The American Naturalist, 134: 344-364.
Rand, A. S. 1964. Ecological distribution in anoline lizards of Puerto Rico. Ecology, 45: 745-752.
Reagan, D. P. 1992. Congeneric species distribution and abundance in a three-dimensional habitat: The rain forest anoles of Puerto Rico. Copeia, 1992: 392-403.
Rodríguez-Schettino, I. (ed.). (1999): The Iguanid Lizards of Cuba. University of Florida Press, Gainesville, 428 pp.
Schoener, T. W. 1977. Competition and the niche. (pp. 35-136). En: Gans, C. y D. W. Tinkle (Eds.), Biology of the Reptilia, vol. 7.
Schoener, T. W. 1983. Population and Community Ecology (233-239). En: Huey, R. B., E. R. Pianka y T. W. Schoener (Eds.), Lizard Ecology. Studies of a model organism. Harvard University Press, Cambridge, 501 pp.
Schwartz, A. y R. W. Henderson. 1991. Amphibians and Reptiles of the West Indies: Descriptions, Distributions and Natural History. University of Florida Press, Gainesville, 720 pp.
Sokal, R. R. y F. J. Rohlf.1979. Biometría. Principios y métodos estadísticos en la investigación biológica. H. Blume Ediciones, Madrid.
Socarrás, E. 1994. Caracterización herpetológica del Bosque Siempre verde Micrófilo en el Área Protegida “Loma del Puerto” de Cayo Coco. En: Segundo Taller de Biodiversidad, Santiago de Cuba. BIOECO, Resúmenes.
Torres López, J., T. M. Rodríguez-Cabrera y R. Marrero Romero. 2017. Reptiles (376-411). En: Mancina, C. A. y D. D. Cruz (Eds.), Diversidad biológica de Cuba: métodos de inventario, monitoreo y colecciones biológicas. Editorial AMA, La Habana, 502 pp.
Williams, E. E. 1972. The origin of faunas. Evolution of lizard congeners in a complex island fauna: a trial analysis. Evolutionary Biology, 6: 47-89.