
INTRODUCTION
Spinycheek sleepers (genus Eleotris) are distributed from North Carolina to Brazil in the Western Atlantic Ocean (Guinarães-Costa et al., 2016). They inhabit mangrove estuaries and freshwater streams and are considered important predators in these habitats (Pezold & Cage, 2001; Guinarães-Costa et al., 2016). Generally the Eleotris adults are too small to be used for human food, but the larvae are a component of “tismiche” and are eaten along with a variety of amphidromous goby larvae (Nordlie, 2012).
There are three eleotrid species known from freshwater habitats on Montserrat. Schmidt and McMullin (2015) collected Gobiomorus dormitor and Eleotris amblyopsis. Subsequent observations on the island have documented Eleotris perniger as more common than E. amblyopsis, which is typical of Caribbean island ichthyofauna (Pezold & Cage, 2001). Eleotris perniger (Cope, 1871), the Smallscaled Spinycheek Sleeper, is considered amphidromous (McDowall, 1997; Nordlie, 2012; Cooney & Kwak, 2013).
Pezold and Cage (2001) delineated the distributions of the three Eleotris species known from the Caribbean and provided data on meristic and morphometric differences among them.
The larval stages of these species, however, remain difficult to distinguish. Larvae of E. perniger are marine, and transforming larvae enter freshwater streams after spending an average of 65.2 days at sea (Engman, 2007). Baldwin and Smith (2003) and Yeung and Ruple (2006) illustrated and described Eleotris larvae from Belize, but could not determine the species. Victor (2016) described and figured E. amblyopsis and E. perniger larvae, which he distinguished by modal pectoral fin ray counts (16 in E. amblyopsis, 18 in E. perniger), but there is substantial overlap in this meristic between the two species (Pezold & Cage, 2001).
OBJECTIVES
To describe newly transformed larvae of Eleotris perniger collected in a freshwater stream.
To discuss the habitat of this species on the island of Montserrat.
MATERIALS AND METHODS
Field collection. Specimens were collected in the freshwater mouth of the Bunkum River, a small stream on the west (Caribbean Sea) side of Montserrat (Fig. 1) on January 16, 2016. Fishes were captured in an “eel mop”, a passive device designed to capture the glass eel stage of American eel, Anguilla rostrata (Silberschneider et al., 2001; Sullivan et al., 2009). Fishes were shaken out of the mop onto a small-mesh net. Small eleotrids were preserved in 10 % formalin except one mostly unpigmented specimen placed in 95 % ethanol for DNA analysis. Specimens were catalogued into the New York State Museum fish collection (NYSM 73740). Other specimens reported here from various locations and dates were taken with small seines.
DNA analysis. Genomic DNA was extracted from a portion of one specimen using a DNeasy Blood and Tissue Kit (Qiagen) and a 666 bp region of the Cytochrome Oxydase I gene was amplified using FishF1 and FishF2 primers designed by Ward et al. (2005). An additional 530 bp region of the 16S gene was also amplified, using primers designed by Palumbi et al. (1991). PCR products were purified using a Qiaquick PCR Purification Kit (QIAGEN) and sequenced using the same primers as the PCR (DNA Analysis Facility on Science Hill, Yale University, New Haven, CT). Complimentary forward and reverse sequences were aligned and edited in BioEdit (Hall, 1999) and the resulting sequence was used to query the nucleotide databases of GenBank (blastn). Returned sequences with a high percentage match to the query sequence, as well as an outgroup sequence, were downloaded and compared to our sequence through a BLAST search.
Meristics and morphometrics. Pezold and Cage (2001) distinguished E.amblyopsis and E. perniger in the Lesser Antilles by lateral line scale counts and the configuration of cephalic neuromasts. However, none of these characters are visible on marine or freshwater larvae. Specimens >14 mm standard length (SL) were identifiable as E.perniger using lateral line scale counts and cephalic neuromast patterns. Meristic data were gathered on those juvenile and adult specimens of E. perniger collected on Montserrat because Pezold and Cage (2001) did not include specimens from Montserrat in their analyses. Counts were done under a binocular dissecting microscope at 10-25X. These data were then compared to specimens from other Caribbean islands. Morphometric data were gathered from three transitional larvae from Montserrat using dividers under a binocular dissecting microscope. Fin ray counts and morphometric data using dividers on marine larvae were gathered from a photograph (Victor, 2016) and an illustration (Baldwin & Smith, 2003). Data for juveniles and adults were taken from Pezold and Cage (2001).
Specimens examined. All specimens were from Montserrat and were catalogued in the New York State Museum (Fig. 1): 72020-Wetland north end of Carr’s Bay, Little Bay Estate, January 6, 2015, 11 specimens, 14-73 mm SL; 73718-Mouth of Bottomless Ghaut, Blakes Estate, January 16, 2016, 3 specimens, 14-15 mm SL; 73738-Mouth of Bottomless Ghaut, Blakes Estate, 1 specimen, 46 mm SL; 73739-Mouth of Bunkum River, St. Peters, January 16, 2016, 1 specimen, 37 mm SL; 73740-Mouth of Bunkum River, St. Peters, January 16, 2016, 5 specimens, 14-15 mm SL; 73755-Mouth of Bottomless Ghaut, Blakes Estate, January 13, 2016, 11 specimens, 13-27 mm SL; 73775-Mouth of Bottomless Ghaut, Blakes Estate, January 14, 2016, 1 specimen, 32 mm SL; 75583-Mouth of Bunkum River, St. Peters, January 8, 2017, 9 specimens, 13-62 mm SL; 75702-Brimm’s Ghaut, Marguerita Bay, town of Overlook, Blakes Estate, January 16, 2017, 6 specimens, 18-26 mm SL; 78427-Brimm’s Ghaut, Marguerita Bay, town of Overlook, Blakes Estate, December 31, 2017, 5 specimens, 20-24 mm SL.
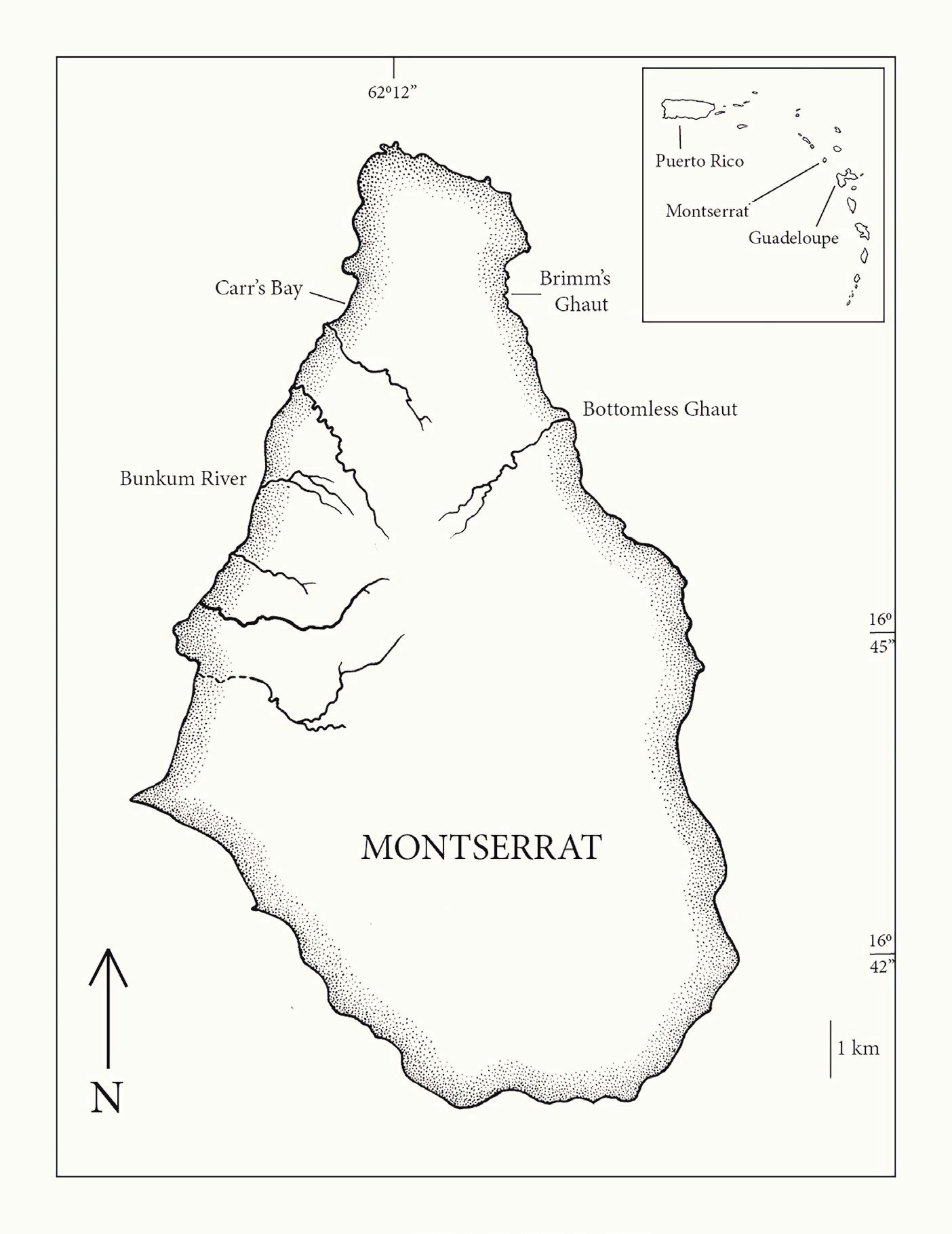
Figure 1. Montserrat, West Indies, showing freshwater streams where Eleotris perniger was collected. Note that all collections of this species were made right on the coastline.
RESULTS
Identification. The COI sample from Montserrat differed by 2 bp (out of 666, 0.3 %) from E. perniger sequenced from Barbados (Guinarães-Costa et al., 2016). The 16S sample from Montserrat differed by 2 bp (out of 530, 0.4 %) from a Genbank sequence of E. perniger (NCBI #KX095207.1). Therefore, the transitional larvae collected on Montserrat are identified as Eleotris perniger. Mean of pectoral fin ray counts was 17.8 with a mode of 18 (Table I) also indicating that the specimens were E. perniger.
Table I. Meristics and morphometrics of Eleotris perniger life stages

Color Pattern (in life). The transitional larvae were generally transparent. Melanophores were aligned along the hypaxial myotomes posterior to the anus, giving the individuals a partially diagonally striped appearance. This observation contrasts with the essentially unpigmented specimens collected in marine waters (Baldwin & Smith, 2003; Victor, 2016).
Color Pattern (in preservative). Preservation in formalin caused melanophores to expand, obscuring the pattern described above (Fig. 2). Large, dark melanophores are distributed along the epaxial and hypaxial myomeres all along the body. These melanophores become more scattered on the ventro-lateral side anterior to the anal fin. The ventro-lateral side is immaculate above and posterior to the anal fin.
There is a very dark, rectangular basicaudal spot that is the same depth as the caudal peduncle. The caudal rays have a dark basal spot, which is separated from the basicaudal spot, and more extensive in the center of the fin than on the dorsal and ventral edges. All other fins including the numerous accessory caudal rays are immaculate or have very tiny basal spots.
The side of the head has scattered melanophores and a dark line extending from the premaxilla through the eye, diverging into two lines posterior to the eye. The spots are much smaller between the postorbital lines and on the nape.
The ventral region is white with a narrow black line extending from the isthmus posteriorly to just beyond the pelvic fins. A dark blotch is present at the anal fin origin and two dark blotches are present on the caudal peduncle posterior to the anus. Streaky melanophores are present on the branchiostegal membranes and small spots are distributed on the lower surface of the head.
The dorsal region is pale behind the eyes and extending to the dorsal fin origin, becoming darker posterior to the dorsal fins. Small melanophores are scattered throughout the dorsal region.

Figure 2. Different states of early young of Eleotris perniger in a freshwater stream mouth on Montserrat. A, transitional larva of E. perniger, 12.5 mm SL. B & C, larger juveniles of E. perniger (13.2 & 14.0 mm SL, respectively) with more extensive pigmentation.All were collected together (NYSM 73740).
DISCUSSION
Both E. amblyopsis and E. perniger are present on Montserrat, which is typical in Caribbean islands (Pezold & Cage, 2001), E. perniger being the more abundant species. The meristics and morphometrics of E. perniger are very similar to specimens collected on other islands. Despite this description, larvae of the two Eleotris species may still not be distinguishable.
Nordlie (2012) discussed threats to the persistence of amphidromous eleotrids in freshwater streams, identifying damming and water withdrawals as a major concern on Caribbean islands and elsewhere. Smith et al. (2008) documented loss of most eleotrid species and a large decline in the abundance of E. perniger in the Espiritu Santo estuary on Puerto Rico due to upstream water extraction (the spinycheeck sleeper was identified as E. pisonis in that study). Eleotris perniger is restricted to low gradient streams or brackish water due to a poor ability to surmount barriers (Kwak et al., 2007).
The populations of E. perniger (and also E. amblyopsis) on Montserrat may be living in a more vulnerable environment than on some of the larger Caribbean islands. Currently the streams on Montserrat are quite small with a high gradient. The Belham River was the largest freshwater stream on the island with a relatively low gradient, but it was buried by 10 m of ash and sand beginning with the 1995 volcanic eruption and by lahars in subsequent years (Barclay et al., 2007). Because of the steep gradient in the remaining streams and the relatively poor ability of these fishes to surmount barriers, the available habitat for Eleotris species can be measured in tens of meters in most streams on Montserrat. We noted risks to fishes associated with that habitat: flows can be intermittent in the dry season, much of the flow can become subsurface near the ocean, and water quality deteriorates due to stagnation under dry conditions. We collected transitional larvae of E. perniger in a stream mouth in mid-January. If residence in the ocean is about 65 days (Engman, 2007), spawning would then occur around mid-November during Montserrat’s rainy season when freshwater stream flow should be more substantial.
An example of how tenuous existence might be for this species comes from observations made during collection of a few small specimens of E. perniger. These observations were made in the Belham Valley in a brackish pool inland of the berm on a beach. There was a pool of water about 10-15 cm deep with a small trickle of freshwater coming from subsurface flow. Many amphidromous organisms were present in the pool, mostly early instars of prawns (Decapoda: Macrobranchium spp.). Water quality was poor, as evidenced by hundreds of dead prawns. We collected several small ( <15 mm SL ) E. perniger by lifting small stones on the pool margins and picking up the individual sleepers. We also collected small juveniles of Awaous banana, an amphidromous gobiid. All of these individual amphidromous organisms had elected to move into the pool, probably during a high surf event, but this pool and all the organisms were gone within two days.
The freshwater streams on Montserrat are too small to use for significant power generation but there are additional negative anthropogenic effects on the streams. Some streams receive sewage effluent and water is being removed from some to supply drinking water to the island.
There is pressure to increase the drinking water withdrawal for bottling and sale off-island.
ACKNOWLEDGEMENTS
Specimens were collected under a Memorandum of Understanding between the Government of Montserrat and Bard College at Simon’s Rock. We appreciate the continued support of E. Corbett and S. Mendes, Montserrat Ministry of the Environment. Chris Bowser helped collect specimens with the eel mop. Kathleen Schmidt drafted Figure 1 and helped collect specimens from the Belham River puddle. Jose Calle helped with Spanish translation.
Literatura Citada
Baldwin, C. C., & D. G. Smith. 2003. Larval Gobiidae (Teleostei: Perciformes) of Carrie Bow Cay, Belize, Central America. Bulletin of Marine Science, 72: 639-674.
Barclay, J., J. Alexander, & J. Sušnik. 2007. Rainfall-induced lahars in the Belham Valley, Montserrat, West Indies. Journal of the Geological Society of London, 164: 815-827.
Cooney, P. B., & T. G. Kwak. 2013. Spatial extent and dynamics of dam impacts on tropical island freshwater fish assemblages. BioScience, 63: 176-190.
Engman, A. C. 2007. Amphidromous fish recruitment and its ecological role in Caribbean freshwater-marine ecotones. Ph.D. Thesis, University of North Carolina, Chapel Hill, North Carolina, USA. 169 pp.
Guinarães-Costa, A., M. Vallinoto, T. Giarrizzo, F. Pezold, H. Schneider, & I. Sampaio. 2016. Molecular evidence of two new species of Eleotris (Gobiiformes: Eleotridae) in the Western Atlantic. Molecular Phylogenetics and Evolution, 98: 52-56.
Hall, T. A. 1999. BioEdit: A user-friendly biological sequence-alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41: 95-98.
Kwak, T. J., P. B. Cooney, & C. H. Brown. 2007. Fishery population and habitat assessment in Puerto Rican streams. Phase I, Final Report. Federal Aid in Sport Fish Restoration Project F-50. U.S. Geological Survey, North Carolina Cooperative Fish and Wildlife Research Unit Department of Biology, North Carolina State University, Chapel Hill, North Carolina, USA. 195 pp.
McDowall, R. M. 1997. The evolution of diadromy in fishes (revisited) and its place in phylogenetic analysis. Reviews in Fish Biology and Fisheries, 7: 443-462.
Nordlie, F. G. 2012. Life-history characteristics of eleotrid fishes of the Western hemisphere, and perils of life in a vanishing environment. Review of Fisheries Biology and Fisheries, 22: 189-224.
Palumbi, S. R., A. Martin, W. O. McMillan, L. Stice, & G. Grabowski. 1991. The simple fool’s guide to PCR, version 2.0. Department of Zoology and Kewalo Marine Laboratory, University of Hawaii, Honolulu, HI, 45 pp.
Pezold, F., & B. Cage. 2001. A review of the spinycheek sleepers, genus Eleotris (Teleostei: Eleotridae), of the Western hemisphere, with comparison to West African species. Tulane Studies in Zoology and Botany, 32: 1-45.
Schmidt, R. E., & E. R. McMullin. 2015. Fresh and brackish-water fishes observed on Montserrat, Lesser Antilles, West Indies. Caribbean Naturalist, 25: 1-12.
Silberschneider, V., B. C. Pease, & D. J. Booth. 2001. A novel artificial habitat collection device for studying resettlement patterns in anguillid glass eels. Journal of Fish Biology, 58: 1359-1370.
Smith, K. L., I. Corujo Flores, & Cm. Pringle. 2008. A comparison of current and historical fish assemblages in a Caribbean island estuary: Conservation value of historical data. Aquatic Conservation of Marine and Freshwater Ecosystems, 18: 993-1004.
Sullivan, M. C., M. J. Wuenschel, & K.W. Able. 2009. Inter and intra-estuary variability in ingress, condition, and settlement of the American eel Anguilla rostrata: Implications for estimating and understanding recruitment. Journal of Fish Biology, 74: 1949-1969.
Victor, B. 2016. A photographic guide to the larvae of coral reef fishes. Suborder Gobioidei. Available at: www.coralreeffish.com (accessed: July 15, 2020).
Ward, R. D., T. S. Zemlak, B. H. Innes, P. R. Last, & P. D. N. Hebert. 2005. DNA barcoding Australia’s fish species. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 360: 1847-1857.
Yeung, C., & D. Ruple. 2006. Eleotridae: Sleepers, pp. 2079-2087. In: Richards, W. J. (Ed) Early Stages of Atlantic Fishes. CRC Press, Boca Raton, FL, 2640 pp.