
INTRODUCCIÓN
Las aves conforman uno de los grupos de vertebrados más diversos del archipiélago cubano. Dentro de este grupo el endemismo alcanza el 7.04 %, con un total de 26 especies, ocupando Cuba un lugar destacado respecto al resto de las islas caribeñas (Raffaelle et al., 1998; Garrido y Kirkconnell, 2011). De acuerdo con el registro paleontológico del Cuaternario, la avifauna cubana incluyó en el pasado un mayor número de endémicos, entre los cuales sobresalían los raptores de las familias Accipitridae, Falconidae, Tytonidae, Strigidae y Teratornithidae, la última extinta globalmente (Arredondo y Arredondo, 2002; Suárez, 2004; Suárez y Olson, 2009; Suárez y Olson, 2015; Orihuela, 2019; Suárez y Olson, 2020a; Suárez, 2020a, b). Al igual que en el resto de Las Antillas, las aves raptoras cumplieron el rol de controladores de las poblaciones de mamíferos, dada la inexistencia de mamíferos carnívoros autóctonos (Arredondo, 1976, 1982). Ese conjunto de aves incluyó especies de gran talla, con hábitos terrestres y vuelo limitado, tal es el caso del búho Ornimegalonyx oteroi, el cual fue de los estrígidos de mayor talla del mundo, con unos 30 kg de peso (Arredondo 1970; Alegre, 2002). Otro ejemplo fue la grulla Antigone cubensis, de alas cortas y una talla que superaba a la grulla cubana actual Antigone canadensis nesiotes, que mide 102 cm del pico a la cola (Garrido y Kirkconnell, 2011). Se han postulado tres eventos de extinción para la avifauna del Caribe durante el Cuaternario (Olson, 1978), el primero aconteció hace ~10 k años, y tuvo implicaciones de carácter global. El siguiente se vincula estrechamente al arribo del hombre aborigen al Caribe hace unos 7 k años, y el tercero a la irrupción y establecimiento de los europeos en el Nuevo Mundo a fines del siglo XV. A pesar de esos eventos, gran parte de la avifauna cubana sobrevivió la etapa aborigen precolombina, alcanzó la era colonial y los siglos XX y XXI. La primera de las especies tratadas es el búho de Arredondo (Pulsatrix arredondoi), extinguido en tiempos prehistóricos, por lo cual se conoce únicamente a partir de restos óseos (Brodkorb, 1968; Orihuela, 2019; Suárez, 2020a). La siguiente es el guacamayo cubano (Ara tricolor), desaparecido en la segunda mitad del siglo XIX (Gundlach, 1876; Navarro, 2015). Y, por último, el carpintero real (Campephilus principalis), que se encuentra en peligro crítico, sin ser observado desde el año 1988 (González y Alayón, 2012).
OBJETIVO
- Dar a conocer nuevos registros paleontológicos y arqueológicos de aves para Cuba, dos de ellas extintas (Ara tricolor, Pulsatrix arredondoi), y una en estado crítico (Campephilus principalis).
MATERIALES Y MÉTODOS
El fragmento de tibiotarso de Ara tricolor fue colectado en el verano de 2006 en el sitio arqueológico A-36 (unidad estratigráfica 54), ubicado en la confluencia de las calles Tacón y Empedrado, en La Habana Vieja. El contexto general del área corresponde a un sector de la Muralla de Mar, obra ingeniera que defendió a partir de 1734 el acceso a la Bahía de La Habana. La unidad estratigráfica 54 se ubica cronológicamente en la primera mitad del siglo XVIII. Se encuentra depositado en la colección arqueozoológica del Gabinete de Arqueología de la Oficina del Historiador de La Habana (CAZGA-265). Por su parte, el pico superior de Ara tricolor se colectó a inicios del año 2011 en el sitio arqueológico A-52, ubicado en áreas del Palacio del Segundo Cabo, sito en calle O´ Reilly esquina a Tacón, también en La Habana Vieja. La pieza se halló en rellenos de basuras expuestos en la cala I (unidad estratigráfica 44), abierta junto a la pared este de la edificación, en el lado próximo al foso del Castillo de la Real Fuerza. De acuerdo con la cultura material exhumada, los estratos de esta cala poseían una cronología que abarcaba todo el siglo XVII. El hueso está depositado en CAZGA-266. Para la determinación taxonómica del fragmento de tibiotarso se utilizó una referencia bibliográfica (Mayr, 2010) y materiales arqueológicos y neontológicos de la colección arqueozoológica del Gabinete de Arqueología de la Oficina del Historiador de La Habana. La determinación taxonómica del pico superior de Ara tricolor fue realizada por el paleornitólogo William Suárez Duque y el primer autor, utilizando como referencias materiales neontológicos de Amazona leucocephala y paleontológicos de Ara tricolor (Wetmore, 1928; Olson y Suárez, 2008b).
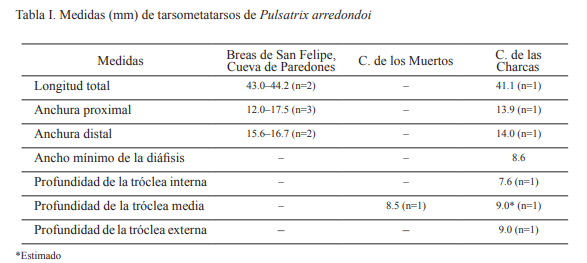
Los dos tarsometatarso de Pulsatrix arredondoi proceden de las Cuevas de los Muertos y de Las Charcas; espeluncas ubicadas en la comunidad Las Charcas, municipio San José de las Lajas, provincia Mayabeque. Ambos ejemplares se colectaron en mayo de 2012 y se encuentran depositados en CAZGA-267 y CAZGA-268. El tarsometatarso de la Cueva de los Muertos se colectó en un contexto arqueológico arcaico. En cambio, el espécimen que procede de la Cueva de Las Charcas corresponde a un contexto paleontológico posiblemente holocénico. La descripción original del taxón (Brodkorb, 1968) fue la fuente bibliográfica utilizada en la identificación de la especie. Las medidas de Pulsatrix arredondoi expuestas en la Tabla I, procedentes de las localidades Breas de San Felipe, provincia Matanzas y Cueva de Paredones, provincia Artemisa, se tomaron de Brodkorb (1968) y Suárez (2005).
El tibiotarso de Campephilus principalis fue colectado el 29 de noviembre de 2013 en la cuadrícula J-4 (unidad estratigráfica 2) del sitio arqueológico arcaico Cueva del Aguacate o de Las Carretas, comunidad Las Charcas, municipio San José de las Lajas, provincia de Mayabeque. Se encuentra depositado en CAZGA-269. Para su identificación se comparó con similares de todos los géneros de la familia Picidae registrados en Cuba (Sphyrapicus, Xiphidiopicus, Melanerpes, Colaptes), adicionalmente se utilizó el artículo de Shufeldt (1900). Las medidas de huesos de C. principalis que aparecen en la tabla 2 proceden de ejemplares de poblaciones de Estados Unidos, y se obtuvieron de dos especímenes depositados en Division of Birds, Smithsonian Institution, National Museum of Natural History (NMNH 291358, macho, colectado por A. T. Wayne, 1890, Everglades, Florida; AMNH 4708, macho, colectado por F. M. Chapman y W. Brewster, 1890, Suwannee River, Florida).
RESULTADOS
Familia Psittacidae
Género Ara Lacépède, 1799
Ara tricolor Bechstein, 1811
Fragmento medio distal de tibiotarso derecho (Fig. 1) y pico superior, ambos corresponden a individuos adultos.
Descripción. La epífisis distal del tibiotarso es ancha mediolateralmente y deprimida en sentido anteroposterior, como ocurre en la familia Psittacidae; en la superficie anterior se observa la ausencia total del puente supratendinal, indicando que representa los géneros Ara o Amazona (Mayr, 2010). En Amazona la diáfisis es algo deprimida anteroposteriormente. En tanto, en el hueso tratado la sección transversal a mitad de la diáfisis es circular. Por otra parte, el cóndilo medial es más alargado longitudinalmente que en Amazona. Asimismo, las dimensiones del tibiotarso superan las de Amazona, como se verá. Aunque al cóndilo medial le falta el borde posterior, se midió su profundidad (6.4 mm) y se estimó la real (6.6 mm); anchura de la diáfisis sobre los cóndilos 6.7 mm; profundidad de la diáfisis sobre los cóndilos 2.5 mm. La anchura del extremo distal, aun estando ausente el cóndilo lateral, es de 8.7 mm, significativamente superior a una muestra de 15 ejemplares actuales de Amazona leucocephala (6.4–7.1 mm). La anchura distal original se estimó en unos 9.0 mm. El pico superior, extraviado al presente, exhibe mayor tamaño y robustez que los de Amazona y Psittacara. Se descarta que la muestra corresponda a una especie exótica de Ara, pues la información documental y arqueológica de los siglos del XVI al XVIII solo alude a sitácidos autóctonos como Ara tricolor, Amazona leucocephala y Psitaccara euops (Torre, 1857; Pérez y Berthe, 1971; Casas, 1987; Jiménez y Arrazcaeta, 2007). Es a partir del siglo XIX que la literatura cubana hace alusión a sitácidos importados desde América continental y África (Pichardo, 1862).
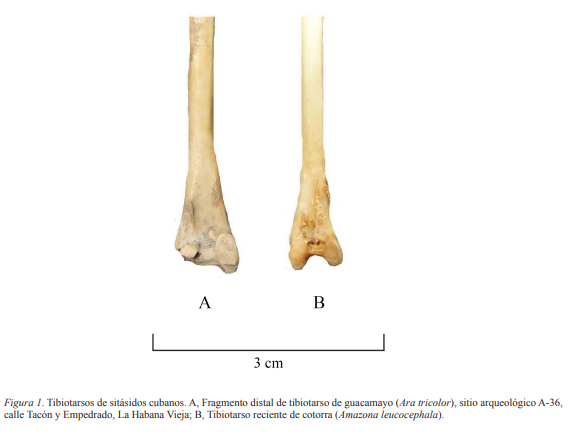
Figura 1. Tibiotarsos de sitásidos cubanos. A, Fragmento distal de tibiotarso de guacamayo (Ara tricolor), sitio arqueológico A-36, calle Tacón y Empedrado, La Habana Vieja; B, Tibiotarso reciente de cotorra (Amazona leucocephala).
Familia Strigidae
Género Pulsatrix Kaup, 1848
Pulsatrix arredondoi, Brodkorb, 1969
Extremo distal de tarsometatarso izquierdo (Fig. 2, A, Tabla I); tarsometatarso izquierdo casi completo (Fig. 2, B); ambos pertenecen a individuos adultos.
Descripción. Tarsometatarso corto y ancho, como en Otus, Bubo, Pulsatrix, Nyctea, Surnia, Glaucidium, Micrathene y Aegolius; mientras que en otros búhos americanos este hueso es largo y estrecho. Tróclea interna tan estrecha como la tróclea media. Foramen distal para la arteria tibialis antiquus prolongado y con una ubicación baja en la diáfisis. Borde interno de la diáfisis casi recto. Difiere de Pulsatrix perspicillata, especie viviente distribuida desde el sureste de México hasta Argentina, por poseer un tarsometatarso más corto y ancho; hipotarso corto, con la muesca proximal más reducida, protuberancia intercotilar y bordes de las cotilas menos elevados (Brodkorb, 1968). Posee puente osificado sobre la acanaladura para el tendón de M. extensor brevis digiti IV. El ejemplar CAZGA-267 presenta dañadas la cresta calcánea del hipotarso y el extremo posterior de la tróclea media. Exhibe color pardo claro y al golpearse emite un sonido cristalino. Corresponde a un individuo adulto, quizás un macho, a juzgar por sus dimensiones (Tabla I), tomado en cuenta que en la familia Strigidae las hembras son mayores que los machos. El ejemplar CAZGA-268, el cual fue sacrificado para datación radiocarbónica de acelerador de masa (14C AMS) que aquí se reporta, era un extremo distal de tarsometatarso izquierdo de un individuo adulto. Este exhibía color amarillento y ligeras evidencias de exposición al fuego, presentando además el extremo posterior de las trócleas interna y medial roto.
Familia Picidae
Género Campephilus Gray, G. R.
1840
Campephilus principalis (Linnaeus, 1758) Diáfisis de tibiotarso izquierdo de un individuo adulto (Fig. 3,
Tabla II). Descripción. Tibiotarso de mayores dimensiones que sus similares de los otros
géneros representados en Cuba: Sphyrapicus, Xiphidiopicus, Melanerpes y Colaptes (Tabla II). Aún faltandole los extermos proximal y distal, se
observa que el tibiotarso es relativamente corto y robusto. Generalmente se considera
que el acortamiento de los huesos incrementa la fuerza afectiva de los músculos
insertados en ellos, en tanto decrese
proporcionalmente la rapidez de su acción. El acortamiento de los huesos de la patas es
una vía mediante la cual se puede incrementar la eficiencia de fijación de los huesos
distales, mientras el ave se acerca al árbol (Spring, 1965). La diáfisis se muestra ligeramente arqueada hacia afuera, o sea, hacia el
borde fibular, y algo deprimida anteroposteriomente. En vista posterior (cara caudal),
una linea recta recorre la diafisis longitudinalmente, conformando una cresta hacia el
extremo proximal. En el borde lateral del hueso se
observa la cresta fibular corta, proximalmente baja, incrementando su altura
gradualmente, hasta su término distal. La conformación de la cresta fibular tiene el efecto de mantener el extremo distal del peroné bien
distante del cuerpo del tibiotarso (Shufeldt, 1900).
DISCUSIÓN
Durante el Cuaternario se produjeron varios eventos de
índole natural y antrópica que causaron la extinción total o parcial de un
importante cúmulo de los vertebrados endémicos del Caribe, entre ellos las aves
(Olson, 1978; Silva et al., 2007;
Steadman y Takano, 2013; Orihuela
et al., 2020a, b). La avifauna fósil de Cuba comprende 34
especies de aves extintas y extirpadas, de ellas 22 endémicas, 12 compartidas
con otras islas caribeñas o con áreas continentales y una de estatus dudoso
(Orihuela, 2019; Suárez, 2020a, b; Suárez y
Olson, 2015, 2020a, b; W. Suárez, com pers., 2020). Entre los grupos más
conspicuos de la avifauna cubana pasada estuvieron los estrígiformes,
representados por búhos y lechuzas de gran talla como Ornimegalonyx
oteroi, Tyto cravesae,
T.
pollens y T. noeli, y
lechuzas de talla pequeña como Tyto maniola
(Suárez y Olson, 2015; Orihuela, 2019; Suárez, 2020a). También hubo un estrígido
de talla menor, se trataba de Pulsatrix arredondoi, el cual se conocía previamente de cuatro localidades:
una en la provincia de Sancti Spiritus, otra en
Matanzas y dos en Artemisa (Brodkorb, 1968; Arredondo y González, 1982; Martínez
y Rives Pantoja, 1990; Suárez, 2005).
En la presente investigación se registran dos nuevas
localidades para P. arredondoi,
ambas situadas en cuevas abiertas en la Llanura Cársica Meridional de la
provincia Mayabeque, particularmente en el municipio San José de las Lajas. La
primera de esas espeluncas se nombra Cueva de Las Charcas. El depósito donde se
colectó el tarsometatarso de P. arredondoi (CAZGA-267) corresponde a un contexto paleontológico del
Cuaternario, conformado por sedimentos rojizos aluviales, el cual se ubicaba por
debajo de una estratificación arqueológica. Dos eventos diacrónicos dieron
origen a este depósito paleontológico:
primero, el transporte mecánico de las aguas y, segundo, los agregados óseos
derivados de egagrópilas generadas por una lechuza extinta, posiblemente
Tyto
noeli. La fauna fósil registrada la componía
una amplia variedad de vertebrados propios de
un clima cálido y húmedo, con bosques, sabanas y cuerpos de agua dulce
estacionales. Estos son: anfibios (Peltophryne sp.),
reptiles (Chelonoidis cubensis, Chilabothrus angulifer), aves (Burhinus bistriatus, Rallus sp.,
Tyto
noeli, Buteogallus borrasi, Falco kurochkini, Cyanolimnas cerverai, Columbina passerina,
Columbidae indet.), mamíferos (Acratocnus
antillensis, Neocnus
gliriformis, Megalocnus rodens,
Parocnus browni, Capromys pilorides, Geocapromys columbianus, Macrocapromys acevedo,
Mysateles prehensilis, Mesocapromys nanus/kraglievichi,
Boromys
torrei, B. offella,
Nesophontes
micrus, quirópteros indet.). La segunda
espelunca se nombra Cueva de los Muertos, y a diferencia de la cueva anterior,
el contexto donde se colectó el fragmento de tarsometatarso de P. arredondoi
(CAZGA-268) es de origen arqueológico, correspondiendo culturalmente a los
aborígenes arcaicos. En el contexto abundaban los restos de dieta de especies
comunes en sitios de tierra adentro, asociados a paisajes de bosques y
cuerpos de agua dulce como arroyos y lagunas.
Las especies representadas fueron: crustáceos (Epilobocera cf. gilmani,
Gecarcinus
sp.), reptiles (Chilabothrus
angulifer, Cubophis cantherigerus), aves (Nesotrochis picapicensis, Mycteria americana, Nyctanassa violacea,
Phalacrocorax
olivaceus,
Accipiter
gundlachii, Corvus nasicus), roedores (Capromys pilorides, Mysateles prehensilis, Geocapromys columbianus, Mesocapromys kraglievichi, Boromys torrei,
Boromys
offella, Solenodon cubanus).
El tarsometatarso de P. arredondoi
(CAZGA-268) de la Cueva de los Muertos mostraba una coloración clara, ligeras
huellas de afectación por el fuego y no estaba mineralizado, por lo cual parecía
corresponder al contexto arqueológico, fechado anteriormente en 1190 ± 40 AP
(cal 690–970 AD; Orihuela et al., 2016). Para esclarecer este asunto se sometió el
tarsometatarso a un fechado C14AMS, el cual arrojó una antigüedad de 1390 ± 30 AP (cal
600–680 AD). Este resultado incluye a P. arredondoi en
el lapso aborigen reconocido para Cuba
(6000–500 años AP), (Cooper, 2007) dando cabida, además, a la posibilidad de que
la especie fuera utilizada como alimento por los indocubanos arcaicos. Esta
“fecha de última presencia” demuestra que especies de vertebrados que se
creyeron extintas milenios atrás
sobrevivieron hasta periodos más recientes (Orihuela, 2019; Orihuela
et al., 2020b). La segunda especie implicada es el carpintero real (Campephilus principalis), el cual se encuentra por primera vez en contextos
paleobiológicos cubanos. El fragmento de tibiotarso
de la especie se halló en la Cueva del Aguacate, espelunca muy próxima a las cuevas de
Las Charcas y los Muertos, donde se registró el búho P. arredondoi. Aunque el
resto se colectó en un contexto arqueológico arcaico, partiendo de su apariencia (mineralización, color pardorojizo, ausencia de marcas de
carnicería y patrón de fragmentación de “hueso seco”), es más probable que corresponda
al yacimiento paleontológico que yace bajo los estratos arqueológicos, formado por
sedimentos rojos aluviales que contenían, además,
restos de vertebrados pequeños como anfibios (anuros indeterminados), reptiles
(Leiocephalus
sp., lagartos indeterminados), aves (especies pequeñas
indeterminadas) y mamíferos (Natalus primus,
Nesophontes
micrus, Boromys offella, Boromys torrei,
Geocapromys
columbianus). Las características tafonómicas de esta
tanatocenosis indican que su origen pudiera estar relacionado a dos procesos: uno pudo
ser la muerte in
situ del ejemplar de C. principalis en el bosque exterior o en el bosque establecido en la gran dolina que sirve de acceso a la
cueva. Luego, el transporte mecánicopluvial trasladó el hueso al recinto subterráneo. El
segundo proceso, que incluiría a la mayor parte de la asociación ósea, fue la actividad
trófica de las lechuzas (Tyto). En la dieta antigua y actual de T. furcata se han registrado tres especies de carpinteros Xiphidiopicus percussus, 23 cm de talla; Melanerpes superciliaris,
28 cm; Colaptes
spp., 33–34 cm (Jiménez et al., 2005; López, 2012; Orihuela et al., 2020a), sin embargo, tratándose de un ave de talla grande como
C. principalis (48 cm), portadora, además, de un poderoso pico, es posible que
haya sido presa de las lechuzas Tyto cravesae y
T. noeli, que fueron los titónidos gigantes con mayor
distribución espacial y abundancia.
El contexto paleontológico de la Cueva del Aguacate no posee
fechado C14, no
obstante, criterios paleoambientales deducidos del origen aluvial de los sedimentos
acumulados en esta cavidad, y en la Cueva de Las Charcas, permiten postular que ambos contextos deben ostentar una edad similar. El
origen aluvial de estos sedimentos, y la ecología de algunas de las especies contenidas
en ellos (Cyanolimnas
cerverai, Rallus sp.), indican un
clima húmedo y cálido, con ambientes de bosques y
sabanas estacionalmente inundadas; la presencia de impregnaciones de óxido de manganeso
sobre los huesos colectados en ambas cuevas indica un ambiente reductor, o sea, seco e
inundado alternadamente. Los sedimentos aluviales fueron transportados a las
cuevas durante una época más húmeda que el presente,
posiblemente acontecida en el transcurso del Holoceno Temprano a Medio 8–3 ka (Curtis
et al., 2001). Se considera que entre 8–7 ka hubo una época más húmeda
que el presente, debido a dos factores: 1- el ascenso
del nivel marino, y como consecuencia, del nivel freático general; 2- la ocurrencia de
un pico de pluviosidad que contribuyó al
empantanamiento de las partes bajas del relieve. La combinación de ambos factores
produjo el desarrollo de ambientes de humedales y
lagunas en las llanuras, donde cocodrilos, tortugas y otros animales acuáticos
convivieron con animales terrestres en casi todo el territorio de Cuba
(Iturralde-Vinent, 2003; Jiménez Vásquez et al., 2013). De las
evidencias anteriores se puede deducir que la
antigüedad de los yacimientos paleontológicos de las Cueva del Aguacate y de Las Charcas
pudiera situarse entre 8–3 ka (Holoceno temprano-medio).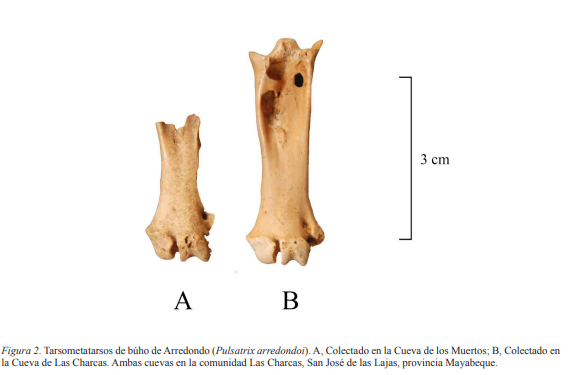
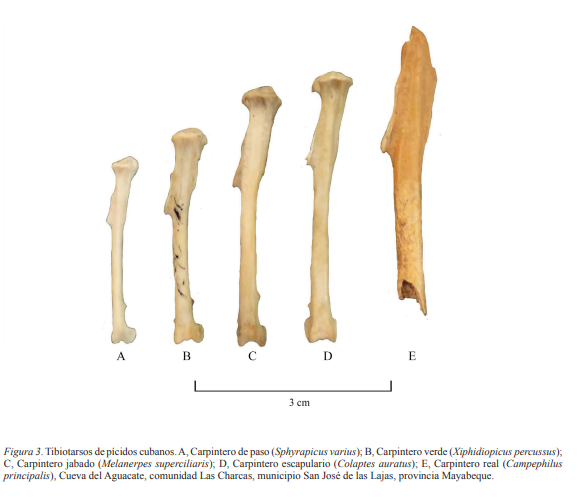

De C. principalis de Cuba se conocen 29 ejemplares disecados, depositados en instituciones cubanas (16) y extranjeras (13). Estos proceden de localidades como Pan de Guajaibón, provincia de Artemisa, Bahía de Cochinos, provincia de Matanzas y montañas de Sagua-Baracoa, en la provincia de Guantánamo (Garrido y Kirkconnell, 2011). Las primeras noticias de la especie datan del siglo XIX, y a fines de ese siglo se consideraba que sus poblaciones iban en decrecimiento debido a la destrucción de los bosques y la caza indiscriminada, sobreviviendo la especie en parajes poco frecuentados por el hombre (Gundlach, 1876). Originalmente habitaba diferentes formaciones de bosques de ciénaga, semideciduo, siempreverde y pinares (Gundlach, 1893). Los últimos avistamientos se realizaron en febrero de 1988 en la localidad de Ojito de Agua, Parque Nacional Alejandro de Humboldt, en la provincia de Guantánamo; en esa ocasión se observaron cinco individuos (González, 2002). La desaparición de los bosques primigenios de Cuba, desde el siglo XVIII (Funes Monzote, 2008), debió ser la causa fundamental de la reducción de sus poblaciones. Consideramos, que el igual que otros grupos zoológicos, la declinación de las poblaciones de C. principalis tuvo un componente climático que inició antes de la llegada del hombre precolombino a Cuba, y otro antrópico, relacionado fundamentalmente con el arribo de los agroceramistas, y luego los europeos en el siglo XV (Orihuela et al., 2020b). Solo la inaccesibilidad de algunas regiones cubanas salvó a Campephilus del ecocidio ocurrido durante la época colonial y neocolonial, y hoy pudieran resguardar los últimos ejemplares.
Como expresaron varios autores (Bond, 1963; Vergara, 2003; Suárez, 2004), las relaciones biogeográficas de la avifauna cubana están dadas, en primer lugar, con la fauna ornítica de Norteamérica. Entre los taxones de indudable origen norteaméricano tenemos a C. principalis. Jackson (2002) consideró que la especie pudo ser introducida en Cuba desde territorio norteamericano a inicios de la era colonial, hipótesis desechada por el registro paleontológico que presentamos. Fleischer et al. (2006) sometieron ejemplares de C. principalis de Cuba y Estados Unidos, procedentes de colecciones, a análisis de ADN antiguo, resultando, en primera instancia, que esta especie y C. imperialis de México conforman un grupo monofilético; por otra parte, se reveló que las especies implicadas distan significativamente entre sí, sugiriendo su posición como especies separadas. Finalmente, los análisis arrojaron que la divergencia entre las especies tratadas ocurrió en el Pleistoceno Medio. Los propios autores consideraron que el Pleistoceno Medio se correspondió con un evento de oscilación global de las temperaturas, que implicó cambios en un plazo de tiempo entre 41–100 ka, significando un periodo de enfriamiento y descenso del nivel marino. La diferencia de nivel respecto al presente fue de más de 30 m. En este lapso se pudo incrementar la extensión de la Península de Yucatán, acortándose la actual distancia de 176 km que la separa de Cuba, favoreciendo este hecho la colonización de nuestro territorio por un carpintero presumiblemente reticente a volar sobre las aguas.
La tercera especie registrada es el guacamayo cubano (Ara tricolor), cuyos restos se hallaron en dos sitios arqueológicos ubicados en el centro histórico de La Habana Vieja, correspondientes cronológicamente a un lapso ubicado entre los siglos XVII y XVIII. Ambos sitios se originaron por el vertimiento de basuras provenientes de la ciudad. La referencia más antigua al guacamayo cubano (A. tricolor) en La Habana Vieja la encontramos en un documento de 1598 (Torre, 1857). En 1688 tenemos otro documento escrito por el viajero italiano Giovanni Gemelli Careri (Pérez y Berthe, 1971), por entonces de visita en la ciudad, donde se alude a la caza de cotorras (A. leucocephala) y guacamayos (A. tricolor) en las proximidades de La Habana. Hasta el momento, la presencia del guacamayo era desconocida en el registro arqueológico de La Habana Vieja, y solo se habían identificado restos de catey (Psittacara euops) en un sitio de la segunda mitad del siglo XVI, y cotorra (A. leucocephala) en varios sitios con fechas de la segunda mitad del siglo XVI a fines del XIX (Jiménez y Arrazcaeta, 2007). La presencia de restos de sitácidos en sitios urbanos habaneros pudiera obedecer a que dichas aves fueron mantenidas en cautiverio o consumidas como alimento, aunque sobre el último aspecto se considera que la carne de la cotorra se puede comer mientras el individuo es joven y la del guacamayo es dura y tiene un olor particularmente desagradable (Sagra, 1845; Gundlach, 1876).
Restos de A. tricolor se registraron en yacimientos paleontológicos del Cuaternario de las provincias de Villa Clara y Cienfuegos (Wetmore, 1928; Arredondo, 1984; Olson y Suárez, 2008b). En estos, la fauna asociada indica que allí hubo ambientes terrestres y cenagosos donde habitaban reptiles Crocodylus rhombifer, Trachemys decussata y aves acuáticas Ciconia lydekkeri, Ardea alba, Tigrisoma mexicanum, Ixobrychus exilis, Anas platyrhynchos, Gallinula chloropus, Cyanolimnas cerverai (Wetmore, 1928; Acevedo y Arredondo, 1982; Arredondo y Arredondo, 2002; Iturralde-Vinent, 2003; Arredondo, 2007; Olson y Suárez, 2008a).
Estos ambientes eran similares al humedal de Zapata, establecido en el sur de la provincia de Matanzas, el cual en el siglo XIX formó parte del área de distribución de A. tricolor (Gundlach, 1876). Del registro paleontológico y las fuentes históricas aludidas, se deduce que los ambientes lacustres, y medios naturales adjuntos, fueron propicios para la vida de los sitácidos. Olson y Suárez (2008b) manifestaron que en A. tricolor la superficie dorsal del cráneo es aplanada, indicando que probablemente consumía semillas muy duras, especialmente de palmas, como también ocurre con las especies grandes de Ara continentales, en sentido opuesto a las especies de talla menor, que son más frugívoras. En el archipiélago cubano las palmas son diversas genéricamente, y es probable que jugaran un rol importante en la dieta del guacamayo cubano, particularmente las especies que habitan en ciénagas, como Sabal, Gastrococus, Copernicia, Coccothrinax, Acoelorraphe, Thrinax y Acrocomia (Acuña, 1964; Oviedo, 2013). Gundlach (1876) testificó que la dieta de la especie cubana consistía en frutos y semillas diversas, entre ellas Palma Real (Roystonea regia).
AGRADECIMIENTOS
Roger Arrazcaeta Delgado, Raúl Mesa Morales, Marcos A. Acosta Mauri, Gabinete de Arqueología, Oficina del Historiador de La Habana (OHH); Jorge A. Garcell Domínguez, Consejo Nacional de Patrimonio Cultural (CNPC); William Suárez Duque, P.O. Box 16477, West Palm Beach, Florida 33165, USA.; S. L. Olson, Megan Spitzer y Christina A. Gebhard, Division of Birds, Smithsonian Institution, Washington, DC, USA; Peter Capainolo, Division of Birds, American Museum of Natural History, New York, USA.
LITERATURA CITADA
Acevedo, M. y O. Arredondo. 1982. Paleozoogeografía y geología del Cuaternario de Cuba: Característica y distribución geográfica de los depósitos con restos de vertebrados. 9na Jornada Científica del Instituto de Geología y Paleontología, La Habana, Resúmenes, 59–70 pp.
Acuña, J. 1964. Flora de las ciénagas de Cuba. Poeyana, 3: 1–10.
Alegre, Y. 2002. Análisis morfofuncional de la locomoción del búho gigante Ornimegalonyx oteroi (Aves: Strigidae) del Cuaternario de Cuba. Tesis de Diploma, facultad de Biología, Universidad de La Habana, 44 pp.
Arredondo, C. 2007. Paleofauna, paleoambiente y subsistencia alimentaria de humanos tempranos en el noroeste de Cuba central. Informe parcial-04 del resultado no. 4 del proyecto PNAP0405 “Reconstrucción paleoclimática de sectores claves de Cuba y el Caribe: Contribución a los estudios de poblamiento y asentamiento de sitios arqueológicos en Cuba, 37 pp.
Arredondo, O. 1970. Nueva especie de ave pleistocénica del orden Accipitriformes (Accipitridae) y nuevo género para las Antillas. Ciencias Biológicas, 4: 1–19.
Arredondo, O. 1976. The great predatory birds of the Pleistocene of Cuba. Smithsonian contribution to the paleobiology, 27: 169–187.
Arredondo, O. 1982. Los Strigiformes fósiles del Pleistoceno cubano. Boletín de la Sociedad Venezolana de Ciencias Naturales, 140: 33–55.
Arredondo, O. 1984. Sinopsis de las aves halladas en depósitos fosilíferos pleisto-holocénicos de Cuba. Instituto de Zoología, Reporte de Investigación, 17: 1–35.
Arredondo, O. y C. Arredondo. 2002. Nueva especie de ave (Falconiformes: Teratornithidae) del Pleistoceno de Cuba. Poeyana, 470–475 (de 1999): 15–21.
Arredondo, O. y N. González. 1982. Nuevo hallazgo de Pulsatrix arredondoi Brodkorb (Aves: Strigidae) del Pleistoceno de Cuba. Miscelánea Zoológica, 16: 1–2.
Bond, J. 1963. Derivation of the Antillean avifauna. Proceedings of Academy of Natural Science, Philadelphia, 115: 79–98.
Brodkorb, P. 1968. An extinct Pleistocene owl from Cuba. Quarterly Journal of the Florida Academy of Science, 31 (2): 112–114.
Casas, B. de las. 1987. Historia de Las Indias. 3 volúmenes. Sociedad Dominicana de Bibliófilos Inc., Editora Corripio, C. por A., Santo Domingo.1560 pp.
Cooper, J. 2007. Registro nacional de arqueología aborigen de Cuba: una discusión de métodos y prácticas. El Caribe Arqueológico, 10: 132–150.
Curtis, J. H., M. Brenner y D. A. Hodell. 2001. Climate change in the Circum-Caribbean (Late Pleistocene to Present) and implications for regional biogeography. En: Woods, C. A. y F. E. Sergile (Eds.), Biogeography of the West Indies: Patterns and Perspectives, 2da edn., CRC Press, pp. 35–54.
Fleischer, R. C., J. J. Kirchman, J. P. Dumbacher, L. Bevier, C. Dove, N. C. Rotzel, S. V. Edwards, M. Lammertink, K. J. Miglia y W. S. Moore. 2006. Mid-Pleistocene divergence of Cuban and North American ivory-billed woodpeckers. Biological Letters, 2: 466–69.
Funes Monzote, R. 2008. From Rainforest to Cane Field: An Environmental History since 1492. The University of North Carolina Press, Chapel Hill, 357 pp.
Garrido, O. H. y A. Kirkconnell. 2011. Aves de Cuba. Comstock Publishing Associates, Cornell University Press, New York, 287 pp.
González, H (ed.). 2002. Aves de Cuba. UPC Print, Vaasa, Finlandia, 162 pp.
González, H. y G. Alayón. 2012. Hoja de datos del taxón, Campephilus principalis bairdii Cassin, 1863. En: González Alonso, H., L. Rodríguez Schettino, A. Rodríguez, C. A. Mancina y I. Ramos García (Eds.), Libro rojo de los vertebrados de Cuba. Editorial Academia, La Habana, 244–246 pp.
Gundlach, J. C. 1876. Contribución a la ornitología cubana. Imprenta La Antilla, La Habana, 364 pp.
Gundlach, J. C. 1893. Ornitología cubana. Imprenta La Moderna, La Habana, 357 pp.
Iturralde-Vinent, M. 2003. Ensayo sobre la paleogeografía del Cuaternario de Cuba. V Congreso de Geología y Minería, La Habana, Memorias Geomin, 54–73 pp.
Jackson, J. A. 2002. Ivory-billed woodpecker (Campephilus principalis). En: Poole, A. y F. Gill, (Eds.), The birds of North America, no. 711. Philadelphia, PA: The Academy of Natural Sciences.
Jiménez, O., M. M. Condis y E. García Cancio. 2005. Vertebrados postglaciales en un residuario fósil de Tyto alba Scopoli (Aves) del occidente de Cuba. Revista Mexicana de Mastozoología, 9: 85–111.
Jiménez, O. y R. Arrazcaeta. 2007. Las aves en la arqueología histórica de la Habana Vieja, Cuba.Gabinete de Arqueología, 7 (7): 17–29.
Jiménez Vázquez, O., L. W. Viñola y A. Sueiro Garra. 2013. Una mirada al pasado de los cocodrilos de Cuba. En: Alonso Tabet, M., R. Ramos Targarona, R. Rodríguez Soberón, J. B. Thorbjarnarson, J. Belliure Ferrer y V. Berovides Álvarez (Eds.), Los Crocodylia de Cuba, Publicaciones Universidad de Alicante, 172–187 pp.
López, Y. 2012. Alimentación de la lechuza (Tyto alba furcata) en Cuba central: presas introducidas y autóctonas. Tesis de Diploma, Facultad de Biología, Universidad de la Habana. 84 pp.
Martínez, A. G. y A. Rives Pantoja. 1990. Cueva Calero: recinto funerario aborigen de Cuba. Revista Cubana de Ciencias Sociales, 24: 142–157.
Mayr, G. 2010. Parrot interrelationships-morphology and the new molecular phylogenies. Emu, 110: 348–357.
Navarro, N. 2015. Endemic birds of Cuba. A comprehensive field guide. Ediciones Nuevos Mundos, 168 pp.
Olson, S. L. 1978. A paleontological perspective of West Indian birds and mammals. Academy of Natural Sciences of Philadelphia, Special Publication, 13: 99–117.
Olson, S. L. y W. Suárez. 2008a. Bare-throated tiger-heron (Tigrisoma mexicanum) from the Pleistocene of Cuba. A new subfamily for the West Indies. Waterbirds, 31 (2): 285–288.
Olson, S. L. y W. Suárez. 2008b. A fossil cranium of the Cuban Macaw Ara tricolor (Aves: Psittacidae) from Villa Clara province, Cuba. Caribbean Journal of Science, 44 (3): 287–290.
Orihuela, J., O. Jiménez Vázquez, y J. F. Garcell. 2016. Modificaciones tafonómicas en restos óseos: ejemplos arqueológicos y paleontológicos de Mayabeque y Matanzas, Cuba. Cuba Arqueológica, 9: 13–36.
Orihuela, J. 2019. An annotated list of Late Quaternary extinct birds of Cuba. Ornitología Neotropical, 30: 57–67.
Orihuela, J., L. Pérez Orozco, J. L. Álvarez Licourt, R. A. Viera Muñoz, y C. Santana Barani. 2020a. Late Holocene land vertebrate fauna from Cueva de los Nesofontes, Western Cuba: Stratigraphy, chronology, diversity, and paleoecology. Palaeontologia Electronica, 23(3):
a57. https://doi.org/10.26879/995
Orihuela, J., L. W. Viñola, O. Jiménez Vázquez, A. Mychajliw, O. Hernández de Lara, L. Lorenzo y J. A. Soto-Centeno. 2020b. Assessing the role of humans on Greater Antillean land vertebrate extinctions: new insights from Cuba. Quaternary Science Reviews, 249 (2020): 106597.
Oviedo, R. 2013. Diversidad vegetal del humedal Ciénaga de Zapata, Matanzas, Cuba. Tesis Doctoral, Universidad de Alicante, 166 pp. + 1 anexo.
Pérez, J. y J. P. Berthe. 1971. La Habana de fines del siglo XVII vista por un italiano. Revista de la Biblioteca Nacional José Martí, 62 (2): 63–85.
Pichardo, E. 1862. Diccionario provincial casi-razonado de vozes cubanas. 3ra edición, Imprenta La Antillana, calle de Cuba núm. 284.
Raffaelle, H., J. Wiley, O. H. Garrido, A. Keith y J. Raffaelle. 1998. A Guide to the Birds of the West Indies. Princeton Univiversity Press, Nueva Jersey, 511 pp.
Sagra, R. de la. 1845. Historia física, política y natural de la Isla de Cuba. Tomo III, establecimiento tipográfico de don Francisco de P. Mellado, Madrid, 220 pp.
Shufeldt, R. W. 1900. On the osteology of the woodpecker. Proceedings of the American Philosophical Society, 39: 578–622.
Silva, G., Suárez Duque, W. y S. Díaz Franco. 2007. Compendio de los mamíferos terrestres autóctonos de Cuba vivientes y extinguidos. Editorial Boloña, La Habana, 465 pp.
Spring, L. W. 1965. Climbing and pecking adaptations in some North American woodpeckers. The Condor, 67: 457–488.
Steadman, D. W. y Takano, O. M. 2013. A late-Holocene bird community from Hispaniola: Refining the chronology of vertebrate extinction in the West Indies. The Holocene, 23 (7) 936–944.
Suárez, W. 2004. Biogeografía de las aves fósiles de Cuba. En: Iturralde-Vinent, M. (Ed.), Origen y evolución del Caribe y sus biotas marinas y terrestres. Centro Nacional de Información Geológica, La Habana.
Suárez, W. S. 2005. La avifauna fósil de “Las Breas de San Felipe”, Martí, Matanzas, Cuba. M.Sc. thesis. Facultad de Biología, Universidad de la Habana, Cuba.
Suárez, W. 2020a. The fossil avifauna of the tar seeps Las Breas de San Felipe, Matanzas, Cuba. Zootaxa, 4780 (1): 1–53.
Suárez, W. 2020b. Remarks on extinct giant owls (Strigidae) from Cuba, with description of a new species of Ornimegalonyx Arredondo. Bulletin of the British Ornithologists’ Club, 140 (4): 382–387.
Suárez W. y S. L. Olson. 2009. A new genus for the Cuban teratorn (Aves: Teratornithidae). Proceedings of the Biological Society of Washington, 122 (1): 103–116.
Suárez, W. y S. L. Olson. 2015. Systematics and distribution of the giant fossil barn owls of the West Indies (Aves: Strigiformes: Tytonidae). Zootaxa, 4020 (3): 533–553.
Suárez, W. y S. L. Olson. 2020a. Systematics and distribution of the living and fossil small barn owls of the West Indies (Aves: Strigiformes: Tytonidae). Zootaxa, 4830: 544–564.
Suárez, W. y S. L. Olson. 2020b. A new fossil vulture (Cathartidae: Cathartes) from Quaternary asphalt and cave deposits in Cuba. Bulletin of the British Ornithologists’ Club, 140 (3): 335–343.
Torre, J. M. de la. 1857. Lo que fuimos y lo que somos o la Habana Antigua y Moderna, Imprenta de Spencer y compañía. O’Reilly, 110, La Habana, 256 pp.
Vergara, R. 2003. Relaciones biogeográficas de la avifauna cubana. II. Biogeografía descriptiva. Ornitología neotropical, 14: 441–467.
Wetmore, A. 1928. Bones of birds from Ciego Montero deposit of Cuba. American Museum Novitates, 301: 1–5.
Cómo citar:Jiménez, O. y Orihuela, J. (2021) «Nuevos hallazgos de aves en contextos paleontológicos y arqueológicos de Cuba», Novitates Caribaea, (17), pp. 163–176. doi: https://www.doi.org/10.33800/nc.vi17.251